CREB - CREB - Wikipedia
CREB-TF (CREB, cAMP жауап элементімен байланысатын ақуыз)[1] ұялы болып табылады транскрипция коэффициенті. Бұл нақты нәрсемен байланыстырады ДНҚ cAMP жауап элементтері (CRE) деп аталатын реттіліктер, осылайша көбейтеді немесе азайтады транскрипция туралы гендер.[2] CREB алғаш рет 1987 жылы а лагері - реттейтін жауап транскрипциясы коэффициенті соматостатин ген.[3]
Транскрипциясы CREB арқылы реттелетін гендерге мыналар жатады: c-fos, BDNF, тирозин гидроксилазы, көптеген нейропептидтер (сияқты соматостатин, энкефалин, VGF, кортикотропинді шығаратын гормон ),[2] және сүтқоректілерге қатысатын гендер тәуліктік сағат (PER1, PER2 ).[4]
CREB құрылымы мен функциясы жағынан тығыз байланысты CREM (cAMP жауап элементінің модуляторы ) және ATF-1 (транскрипция коэффициентін белсендіру-1 ) белоктар. CREB ақуыздары көптеген жануарларда, соның ішінде адамдарда да болады.
CREB-де жақсы құжатталған рөл бар нейрондық пластика және ұзақ мерзімді жад мидағы қалыптасу және түзілуінде ажырамас болып көрсетілген кеңістіктік жады.[5] CREB регуляциясы патологияға байланысты Альцгеймер ауруы және CREB экспрессиясын арттыру Альцгеймер ауруы үшін мүмкін терапевтік мақсат ретінде қарастырылады.[6] CREB-тің де рөлі бар фототренинг сүтқоректілерде.
Кіші типтер
Келесі гендер CREB немесе CREB тәрізді ақуыздарды кодтайды:
- CREB1 (CREB1 )
- CREB2 атауы өзгертілді ATF4 (ATF4 )
- CREB3 (CREB3 )
- CREB5 (CREB5 )
- CREB3L1 (CREB3L1 )
- CREB3L2 (CREB3L2 )
- CREB3L3 (CREB3L3 )
- CREB3L4 (CREB3L4 )
Құрылым
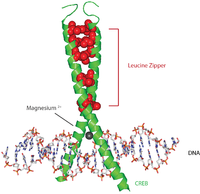
CREB ақуыздары әртүрлі киназалардан фосфорлану арқылы белсендіріледі, соның ішінде PKA, және Ca2+/ кальмодулинге тәуелді протеинкиназалар Serine 133 қалдықтарында.[7] Белсендірілген кезде CREB ақуызы CRE promoter 5 ’ағыны аймағына қосылу үшін басқа транскрипциялық коактиваторларды қабылдайды. Гидрофобты лейцин аминқышқылдары альфа спиралінің ішкі шеті бойында орналасқан. Бұл лейцин қалдықтары димер түзетін басқа CREB ақуызының лейцин қалдықтарымен тығыз байланысады. Бұл лейцин қалдықтарының тізбегі лейциндік сыдырма мотиві. Ақуыздың құрамында ДНҚ-мен байланысуды жеңілдететін магний ионы бар.
cAMP жауап элементі
The cAMP жауап элементі (CRE) - бұл жауап элементі құрамында 5'-TGACGTCA-3 ’жоғары сақталған нуклеотидтер тізбегі бар CREB үшін. CRE сайттары әдетте гендердің жоғарғы жағында орналасқан промоутер немесе күшейткіш аймақтар.[8] Адам геномында шамамен 750 000 палиндромды және жартылай орналасқан CRE бар. Алайда бұл сайттардың көпшілігі цитозиннің әсерінен байланыссыз қалады метилдену, бұл ақуыздармен байланысуға физикалық кедергі келтіреді.[9]
Қимыл механизмі
Оқиғалардың типтік (біршама жеңілдетілген болса да) тізбегі келесідей: сигнал ұяшық бетіне келіп, сәйкес рецепторды іске қосады, бұл а шығаруға әкеледі екінші хабаршы мысалы, cAMP немесе Ca2+, ол өз кезегінде а ақуыз киназасы. Бұл протеин киназасы трансляцияланады жасуша ядросы, онда ол CREB ақуызын белсендіреді. Белсендірілген CREB ақуызы CRE аймағымен байланысады, содан кейін онымен байланысады CBP (CREB-байланыстыратын ақуыз), оны белгілі бір гендерді қосуға немесе өшіруге мүмкіндік беретін оны активтендіреді. CREB-дің ДНҚ-мен байланысуы оның негізгі лейциндік найзағай домені арқылы жүзеге асырылады (bZIP домені ) суретте көрсетілгендей.
Мидың қызметі
CREB әр түрлі органдарда көптеген функцияларды атқарады және оның кейбір функциялары миға байланысты зерттелген.[10] Ішіндегі CREB ақуыздары нейрондар ұзақ мерзімді естеліктерді қалыптастыруға қатысады деп ойлайды;[11] бұл теңіз ұлуында көрсетілген Аплизия, жеміс шыбыны Дрозофила меланогастері, жылы егеуқұйрықтар және тышқандарда (қараңыз. қараңыз) Молекулалық және жасушалық танымдағы CREB ).[1] CREB кеш кезеңі үшін қажет ұзақ мерзімді потенциал. CREB дамуында да маңызды рөл атқарады нашақорлық және одан да көп психологиялық тәуелділік.[12][13][14] CREB активаторы және репрессорлық формалары бар. CREB белсенді емес түрін шамадан тыс әсер ету үшін генетикалық инженерияланған шыбындар ұзақ мерзімді есте сақтау қабілетін жоғалтады. CREB мидағы CREB және CREM жойылған генетикалық инженерлік тышқандарда көрсетілгендей, нейрондардың өмір сүруі үшін де маңызды. Егер CREB бүкіл дамып келе жатқан тышқан эмбрионында жоғалып кетсе, тышқандар туылғаннан кейін бірден өледі, бұл тағы да CREB-тің нейрондық тіршілік етудегі маңызды рөлін көрсетеді.
Аурудың байланысы
Мидағы CREB қызметінің бұзылуы оның дамуына және дамуына ықпал етуі мүмкін Хантингтон ауруы.
CREB KID доменімен әрекеттесетін ақуыздың ауытқулары, CREB байланыстыратын ақуыз, (CBP) байланысты Рубинштейн-Тайби синдромы.
CREB-нің жеткіліксіз жұмыс істеуі байланысты екендігі туралы бірнеше дәлел бар негізгі депрессиялық бұзылыс.[15] CREB шамадан тыс экспрессиясы бар депрессиялық егеуқұйрықтар тісжегі гирусы антидепрессанттармен өңделген егеуқұйрықтарға ұқсас әрекет етті.[16] Өлгеннен кейін жүргізілген зерттеулерден емделмеген негізгі депрессиялық бұзылыстары бар науқастардың кортекстері сау бақылаулармен де, антидепрессанттармен емделген пациенттермен салыстырғанда CREB концентрациясы төмендейтіндігі дәлелденді.[16] CREB функциясын байланыстыру нәтижесінде пайда болатын сигнал беру жолы арқылы модуляциялауға болады серотонин және норадреналин постсинаптикалық G-ақуызды байланысқан рецепторларға. Осы нейротрансмиттердің дисфункциясы үлкен депрессиялық бұзылуларға да қатысты.[15]
CREB сонымен қатар қатерлі ісіктің кейбір түрлерінің өсуіне қатысады деп есептеледі.
Циркадтық ырғақтарға қатысу
Қызығушылық сүтқоректілердің тәуліктік сағаты жарық индукциясы арқылы орнатылған БІР. Жарық құрамында меланопсин бар жарық сезгіш торлы ганглион жасушалары сигнал беретін супрахиазматикалық ядро (SCN) арқылы ретиногипоталамикалық тракт (RHT). RHT қозуы глутаматтың бөлінуін білдіреді NMDA рецепторлары SCN-де, нәтижесінде SCN-ге кальций ағыны әкеледі. Кальций Ca белсенділігін индукциялайды2+/кальмодулинге тәуелді ақуыз киназалары, нәтижесінде активтендіру PKA, PKC, және CK2.[17] Содан кейін бұл киназалар гендердің экспрессиясын әрі қарай реттейтін тәуліктік түрде CREB фосфорилаттайды.[18] Фосфорланған CREB cAMP жауап элементін таниды және үшін транскрипция факторы ретінде қызмет етеді Per1 және Per2, сүтқоректілердің циркадтық сағатын реттейтін екі ген. Бұл PER ақуызының индукциясы тәуліктік сағатты ашық / қараңғы циклдарға итермелеуі мүмкін, транскрипция-трансляция кері байланыс циклі арқылы циклдық сағатты алға немесе кешіктіре алады. Алайда PER1 және PER2 ақуыздарының индукциясының реакциясы субъективті түнде ғана маңызды.[4]
CREB-тің циркадиандық ырғаққа қатысуының ашылуы
Майкл Гринберг алғаш рет 1993 жылы сүтқоректілердің циркадтық сағатындағы CREB рөлін CREB фосфорлануымен фазалық спецификалық жарық импульстарын корреляциялайтын бірқатар тәжірибелер арқылы көрсетті. Түнде субъективті түнде жарық CREB ақуыз деңгейіне қарағанда CREB фосфорлануын арттырды. In vivo, субъективті түнде фазалық ығысуды тудыратын жарық импульстері SCN-де CREB фосфорлануымен корреляцияланған.[19] 2002 жылы Гюнтер Шутцтің тәжірибелері көрсеткендей, Ser142 фосфорлану орны жоқ мутантты тышқандар жарық импульсіне жауап ретінде mPer1 сағатты реттейтін генді шақыра алмады. Сонымен қатар, бұл мутантты тышқандар ашық қараңғы циклдарға бейімделе алмады.[20]
Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ а б Бурчуладзе; т.б. (1994). «CAMP-жауап беретін элементті байланыстыратын ақуыздың мақсатты мутациясы бар тышқандардағы жетіспейтін ұзақ мерзімді жады». Ұяшық. 79 (1): 59–68. дои:10.1016/0092-8674(94)90400-6. PMID 7923378. S2CID 17250247.
- ^ а б Первс, Дейл; Джордж Дж. Августин; Дэвид Фицпатрик; Уильям С. Холл; Энтони-Сэмюэл Ламантиа; Джеймс О. Макнамара және Леонард Э. Уайт (2008). Неврология (4-ші басылым). Sinauer Associates. 170-6 бет. ISBN 978-0-87893-697-7.
- ^ Монмини, МР; Билезикджян, Л.М. (1987). «Ядролық ақуыздың соматостатин генінің циклдік-АМФ жауап элементімен байланысуы». Табиғат. 328 (6126): 175–178. Бибкод:1987 ж. 328..175М. дои:10.1038 / 328175a0. PMID 2885756. S2CID 4345292.
- ^ а б Дибнер, Чарна; Шиблер, Уели; Альбрехт, Урс (2010). «Сүтқоректілердің күндізгі уақыт жүйесі: орталық және перифериялық сағаттарды ұйымдастыру және үйлестіру» (PDF). Физиологияның жылдық шолуы. 72 (1): 517–549. дои:10.1146 / annurev-physiol-021909-135821. PMID 20148687.
- ^ Сильва; т.б. (1998). «CREB және жады» (PDF). Неврологияның жылдық шолуы. 21: 127–148. дои:10.1146 / annurev.neuro.21.1.127. PMID 9530494. Архивтелген түпнұсқа (PDF) 2008 жылғы 28 тамызда. Алынған 22 қаңтар 2010.
- ^ Альцгеймер миында және Ab емделген егеуқұйрық гиппокампальді нейронында CREB экспрессиясының төмендеуі
- ^ Шейвиц, Адам Дж .; Гринберг, Майкл Э. (1999). «CREB: Жасушадан тыс сигналдардың әр түрлі массивімен белсендірілген ынталандырылған транскрипция факторы». Биохимияның жылдық шолуы. 68 (1): 821–861. дои:10.1146 / annurev.biochem.68.1.821. PMID 10872467.
- ^ Карлезон, АҚШ; Думан, RS; Nestler, EJ (тамыз 2005). «CREB-тің көптеген тұлғалары». Неврология ғылымдарының тенденциялары. 28 (8): 436–445. дои:10.1016 / j.tins.2005.06.005. PMID 15982754. S2CID 6480593.
- ^ Алтареджос, Джудит Ю .; Монтини, Марк (наурыз 2011). «CREB және CRTC ко-активаторлары: гормоналды және метаболикалық сигналдарға арналған датчиктер». Молекулалық жасуша биологиясының табиғаты туралы шолулар. 12 (3): 141–151. дои:10.1038 / nrm3072. ISSN 1471-0072. PMC 4324555. PMID 21346730.
- ^ Carlezon WA, Duman RS, Nestler EJ (тамыз 2005). «CREB-тің көптеген тұлғалары». Неврология ғылымдарының тенденциялары. 28 (8): 436–45. дои:10.1016 / j.tins.2005.06.005. PMID 15982754. S2CID 6480593.
- ^ Кандел, Эрик Р. (14 мамыр 2012). «Естің молекулалық биологиясы: cAMP, PKA, CRE, CREB-1, CREB-2 және CPEB». Молекулалық ми. 5: 14. дои:10.1186/1756-6606-5-14. ISSN 1756-6606. PMC 3514210. PMID 22583753.
- ^ Nazarian A, Sun WL, Zhou L, Kemen LM, Jenab S, Quinones-Jenab V (сәуір 2009). «ПКА және CREB ақуыздарындағы базальды және кокаин тудыратын өзгерістердегі жыныстық айырмашылықтар». Психофармакология. 203 (3): 641–50. дои:10.1007 / s00213-008-1411-5. PMID 19052730. S2CID 24064950.
- ^ Ванг Й, Гезци А, Инь Дж.К., Аткинсон Н.С. (маусым 2009). «BK каналы генінің экспрессиясының CREB реттелуі тез есірткіге төзімділік негізінде жатыр». Гендер, ми және мінез-құлық. 8 (4): 369–76. дои:10.1111 / j.1601-183X.2009.00479.x. PMC 2796570. PMID 19243452.
- ^ DiRocco DP, Scheiner ZS, Sindreu CB, Chan GC, Storm DR (ақпан 2009). «Кокаинді сенсибилизациялауда кальмодулинмен ынталандырылған аденилил циклазаларының рөлі». Неврология журналы. 29 (8): 2393–403. дои:10.1523 / JNEUROSCI.4356-08.2009 ж. PMC 2678191. PMID 19244515.
- ^ а б Белмакер, Р. Х .; Агам, Галила (2008). «Негізгі депрессиялық бұзылыс». Жаңа Англия Медицина журналы. 358 (1): 55–68. дои:10.1056 / nejmra073096. PMID 18172175.
- ^ а б Blendy, JA (2006). «Депрессия мен антидепрессантты емдеудегі CREB рөлі». Биол психиатриясы. 59 (12): 1144–50. дои:10.1016 / j.biopsych.2005.11.003. PMID 16457782. S2CID 20918484.
- ^ Айер, Раджашекар; Ван, Тунфэй; Джилетт, Марта (19 қыркүйек 2014). «Нейрондық функционалдылықтың тәуліктік қақпағы: итерациялық метапластиканың негізі». Жүйелік неврологиядағы шекаралар. 8: 164. дои:10.3389 / fnsys.2014.00164. PMC 4168688. PMID 25285070.
- ^ Обриетан, Карл; Импей, Сорен; Смит, Дэйв; Афос, Хайме; Storm, Derrick R. (11 сәуір 2002). «Супрахиазматикалық ядролардағы CAMP реакциясы элементі-гендік экспрессияның тәуліктік реттелуі». Нейрон. 274 (25): 17748–17756. дои:10.1074 / jbc.274.25.17748. PMID 10364217.
- ^ Гинти, Д.Д .; Корнгаузер, Дж. М .; Томпсон, М. А .; Бадинг, Х .; Мэйо, К.Е .; Такахаси, Дж. С .; Greenberg, M. E. (9 сәуір 1993). «Супрахиазматикалық ядродағы CREB фосфорлануын жарықпен және тәуліктік сағатпен реттеу». Ғылым. 260 (5105): 238–241. Бибкод:1993Sci ... 260..238G. дои:10.1126 / ғылым.8097062. ISSN 0036-8075. PMID 8097062.
- ^ Гау, Даниел; Лембергер, Томас; фон Галл, Шарлотта; Кретц, Оливер; Ле Минь, Нгует; Гасс, Питер; Шмид, Вольфганг; Шиблер, Уели; Корф, Хорст В. (11 сәуір 2002). «CREB Ser142 фосфорлануы циркадиан сағатының жеңіл индукцияланған фазалық жылжуын реттейді». Нейрон. 34 (2): 245–253. дои:10.1016 / S0896-6273 (02) 00656-6. PMID 11970866. S2CID 14507897.
- Библиография
- Лорен Слейтер (2005). Скиннердің қорабын ашу: ХХ ғасырдың керемет психологиялық эксперименттері. Нью-Йорк: W. W. Norton & Company. ISBN 978-0-393-32655-0.
- Barco A, Bailey C, Kandel E (2006). «Айқын және жасырын жадыдағы жалпы молекулалық механизмдер». Дж.Нейрохим. 97 (6): 1520–33. дои:10.1111 / j.1471-4159.2006.03870.x. PMID 16805766.
- Конкрайт М, Монмини М (2005). «CREB: қатерлі ісік ауруын алдын-ала жоспарлаушы». Трендтер Жасуша Биол. 15 (9): 457–9. дои:10.1016 / j.tcb.2005.07.007. PMID 16084096.
- Мантамадиотис Т, Лембергер Т, Блэкманн С, Керн Х, Кретц О, Мартин Вилльба А, Тронше Ф, Келлендонк С, Гау Д, Капфхаммер Дж, Отто С, Шмид В, Шицц Г (2002). «Мидың CREB функциясының бұзылуы нейродегенерацияға әкеледі». Нат. Генет. 31 (1): 47–54. дои:10.1038 / ng882. PMID 11967539. S2CID 22014116.
- Mayr B, Montminy M (2001). «Фосфорлануға тәуелді фактор CREB арқылы транскрипциялық реттеу». Нат. Аян Мол. Жасуша Биол. 2 (8): 599–609. дои:10.1038/35085068. PMID 11483993. S2CID 1056720.
- Инь Дж, Дель Векчио М, Чжоу Х, Тулли Т (1995). «CREB жад модуляторы ретінде: dCREB2 активаторының изоформасының индукцияланған өрнегі дрозофилада ұзақ мерзімді жадыны жақсартады». Ұяшық. 81 (1): 107–15. дои:10.1016/0092-8674(95)90375-5. PMID 7720066. S2CID 15863948.
- Инь Дж, Уоллах Дж, Дель Векчио М, Уайлдер Е, Чжоу Х, Куинн В, Тулли Т (1994). «CREB трансгенінің басым негативті индукциясы дрозофиладағы ұзақ мерзімді жадты арнайы блоктайды». Ұяшық. 79 (1): 49–58. дои:10.1016/0092-8674(94)90399-9. PMID 7923376. S2CID 33623585.
Сыртқы сілтемелер
- http://www.ebi.ac.uk/interpro/entry/IPR001630
- Johannessen, M., Pedersen Delghandi, M., and Moens, U. (2004) - CREB нені қосады? - ұялы сигнал.; 10: 1211-1227. https://web.archive.org/web/20070928090058/http://www.sigtrans.org/publications/what-turns-creb-on/
- https://web.archive.org/web/20060902183214/http://focus.hms.harvard.edu//2001/Oct26_2001/neuroscience.html
- CREB + ақуыз АҚШ ұлттық медицина кітапханасында Медициналық тақырып айдарлары (MeSH)
- Дрозофила Циклді-АМФ жауап элементі А ақуызын байланыстырады - Интерактивті ұшу
- Дрозофила 17A температурада В белогын байланыстыратын циклді-AMP жауап элементі - Интерактивті ұшу