Haplogroup L2 (mtDNA) - Haplogroup L2 (mtDNA)
| Haplogroup L2 | |
|---|---|
| Шығу мүмкін уақыты | 80,000–111,100 YBP[1] |
| Ықтимал шығу орны | Батыс Африка[2] немесе Орталық Африка |
| Арғы ата | L2─6 |
| Ұрпақтар | L2a─d, L2e |
| Мутацияны анықтау | 146, 150, 152, 2416, 8206, 9221, 10115, 13590, 16311!, 16390[3] |
Haplogroup L2 Бұл адамның митохондриялық ДНҚ-сы (mtDNA) гаплотоп, кең таралған заманауи таралуы, әсіресе Субэкваторлық Африкада. Оның L2a қосалқы класы - континенттегі, сондай-ақ афроамерикандықтар арасында жиі және кең таралған mtDNA кластері.
Шығу тегі
L2 - Африкада кең таралған тұқым. Ол 87000 мен 107000 жыл бұрын дамыған деп санайды[4] немесе шамамен. 90,000 YBP.[1] Оның жасы, континент бойынша кең таралуы мен әртүрлілігі Африкада нақты шығу нүктесін сенімді түрде іздеуді қиындатады.[5] Гвинеялықтарда байқалған бірнеше L2 гаплотиптері және басқалары Батыс Африка популяциялар генетикалық сәйкестіктермен бөлісті Шығыс Африка және Солтүстік Африка.[6] Батыс немесе Орталық Африкада L2b, L2c, L2d және L2e шығу тегі ықтимал.[5] L2-дің ерте әртүрлілігін бүкіл Африка континентінде байқауға болады, бірақ төмендегі Subclades бөлімінен көріп отырғанымыздай, ең жоғары әртүрлілік Батыс Африка. Субкладтардың көпшілігі негізінен Батыс және Батыс-Орталық Африкада орналасқан.[7]
2015 жылғы зерттеуге сәйкес, «нәтижелер көрсеткендей, Оңтүстік Африкадағы шығыс желілері Батыс / Орталық Африка шежірелерімен жақында уақыт шкаласында, ал шығыс шежірелер едәуір ежелгі болып көрінеді. Орталық африкалық қайнар көзден шыққан үш сәт байланысты L2: 70-50 ка бір шығыс немесе оңтүстік Африкаға көшу, жалғаннан кейінгі қозғалыс 15–10 ка шығыс африка; оңтүстікке қарай банту кеңеюі, соңғы 5 ка. Комплементарлы популяция және L0a филогеографиялық талдауларда mtDNA генінің айқын дәлелі жоқ. Кейінгі қозғалыс кезінде шығыс және оңтүстік популяциялар арасындағы ағын, бұл шығыс африкалық популяциялар мен банту мигранттары арасындағы қоспаның төмендігі туралы айтады.Бұл, ең болмағанда, алғашқы кезеңдерде банту экспансиясы жергілікті популяциялардың аз қосылуымен демикалық диффузия болғандығын білдіреді ».[8]
Тарату
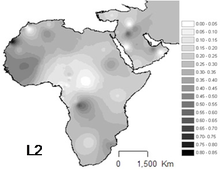
L2 Африкадағы ең көп таралған гаплогруппа болып табылады және ол бүкіл континентте байқалды. Бұл африкалықтардың шамамен үштен бірінде және олардың кейінгі ұрпақтарында кездеседі.
Ең жоғары жиілік олардың арасында болады Мбути пигмийлері (64%).[9] Маңызды қатысу Батыс Африка, әсіресе Сенегал (43-54%).[6] Банту емес популяцияларында да маңызды Шығыс Африка (44%),[10] жылы Судан және Мозамбик.
Бұл әсіресе мол Чад және Канембоу (Үлгінің 38%), бірақ сонымен қатар салыстырмалы түрде жиі кездеседі Көшпелі Арабтар (33%) [Cerny et al. 2007] [5] және Ақан халқы (~33%)[11]
Субкладтар
|
L2-де бес негізгі субаплогруппа бар: L2a, L2b, L2c, L2d және L2e. Осы тұқымдардың ішінен ең көп таралған субклади - L2a, ол Африкада да, Африкада да кездеседі Левант.
L2 Haplogroup тобы арал зиратындағы үлгілер арасында байқалды Кулубнарти, Судан, қай күннен бастап Ерте христиан кезең (AD 550–800).[12]
Haplogroup L2a
L2a кең таралған Африка және ең көп таралған және кең таралған Сахаралық Африканың Гаплогруппасы, сонымен қатар 19% -да жиі кездеседі. Америка африкалықтардың ұрпақтары арасында (Салас және басқалар, 2002). L2a-дің шығу мерзімі шамамен бар. 48,000 YBP.[1]
Бұл әсіресе мол Чад (Үлгінің 38%; арасында 33% дифференциалданбаған L2 Чад арабтары,[13]Банту емес популяцияларында Шығыс Африка (Кения, Уганда және Танзания ) 38% -да.[10] Шамамен 33% Мозамбик[14] және 32% Гана.[11]
Бұл субклайд 2789, 7175, 7274, 7771, 11914, 13803, 14566 және 16294 мутацияларымен сипатталады. Ол L2 жиынтығының 52% құрайды және бүкіл Африкада кең таралған L2 субкледі болып табылады.[15]
L2a мен әртүрліліктің кең таралуы географиялық бастауларды анықтауды қиындатады. Негізгі басқатырғыш - бұл барлық жерде кездесетін Haplogroup L2a, ол Шығыс пен Батысты сол бойына таратуы мүмкін. Сахел Дәліз Солтүстік Африка кейін Соңғы мұздық максимумы немесе бұл кеңеюдің бастауы ертеректе, кейінгі тас дәуірінің басында lie 40,000 жыл бұрын болуы мүмкін.[5][15]
Жылы Шығыс Африка L2a 15% -дан табылды Ніл алқабы –Нубия, 5% Мысырлықтар, 14% Кушит сөйлеушілер, 15% Семит Амхара халқы, 10% Гураг, 6% Тиграй-Тигриния халқы, 13% Эфиопиялықтар және 5% Йемендіктер.[14]
L2a Haplogroup тобы да пайда болады Солтүстік Африка, ең жоғары жиілікпен 20% Туарег, Фулани (14%). Кейбіреулерінің арасында да кездеседі Алжир Арабтар, бұл 10% -да кездеседі Марокко Арабтар, кейбір Марокко Берберлері және Тунис Берберлер. (Уотсон 1997) және т.б., (қырағы 1991) және т.б. 1991 ж.
Препарат берілген науқастарда ставудин емдеу АҚТҚ, Haplogroup L2a төмен ықтималдығымен байланысты перифериялық невропатия жанама әсер ретінде.[16]
L2a1 хаплогруппасы
L2a-ны L2a1-ге бөлуге болады, бұл 16309 ж. Ауысуын сақтайды (Salas және басқалар. 2002).
Бұл субклайд Батыс Африкада әртүрлі жиілікте байқалады Малинке, Wolof, және басқалар; арасында Солтүстік африкалықтар;ішінде Сахел арасында Хауса, Фульбе, және басқалар; жылы Орталық Африка арасында Бамилеке, Фали және басқалары; жылы Оңтүстік Африка арасында Хойсан отбасы, оның ішінде Хве және Банту спикерлер; және Шығыс Африка арасында Кикую бастап Кения.
Барлық L2 қабаттары бар Эфиопия негізінен екі подкладтан, L2a1 және L2b алынған. L2a1 12693, 15784 және 16309 мутацияларымен анықталады. Эфиопиялық L2a1 тізбектерінің көпшілігі 16189 және 16309 нүктелерінде мутацияны бөліседі. Алайда, олардың көпшілігі (33-тен 26) Афроамерикалықтар Share Haplogroup L2a толық тізбектерін L2a1e-3495, L2a1a-3918, L2a1f-5581 және L2a1i-15229 нүктелерінде алмастырулар арқылы төрт подкладқа бөлуге болады. Эфиопиялық 16309 L2a1 үлгілерінде бұл реттіліктің ешқайсысы байқалған жоқ. (Салас 2002) және басқалар.
L2a1 хаплогруппасы да байқалды Махра (4.6%).[17]
Apаплогруппасы L2a1 ежелгі қазба қалдықтарынан табылған Керамикаға дейінгі неолит мәдениет Халулаға айтыңыз, Сирия.[18] Кезінде қазылған үлгі Саванна пасторальдық неолит сайты Люксманда жылы Танзания сонымен қатар L2a1 кладын алып жүрді Қоспаларды кластерлеу анализі бұдан әрі жеке адамның ежелгі Леванттан шыққанын және Саванна Пасторальдық Неолит пен Керамикаға дейінгі Неолит жасаушылары арасындағы тектік байланысты растайтындығын көрсетті.[19]
L2a1a хаплогруппасы
L2a1a подкладиі 3918, 5285, 15244 және 15629-дағы алмастырулармен анықталады. Африканың оңтүстік-шығысында L2a1a және L2a1b екі L2a кластері бар, олардың екеуі де HVS-I позицияларында ауысулармен анықталған. Бұл екеуі де Батыс Африкадан шыққан көрінеді Солтүстік Батыс Африка (сәйкес келетін немесе көршілес түрлердің таралуы көрсеткендей) және Оңтүстік-Африкада немесе қазіргі Оңтүстік-Шығыс Африкаға дейінгі ата-баба популяциясында қатты экспансияға ұшырады.
L2a1a және L2a2 подкладтарындағы соңғы жұлдыздардың жарылуы Бантудың кеңеюіне қол қояды, өйткені Перейра және басқалар ұсынған. (2001).
L2a1a 16286 жылы мутациямен анықталады. L2a1a негізін қалаушы кандидат 2700 (SE 1200) жыл бұрын пайда болды. (Перейра және басқалар 2001). Алайда, L2a1a, (np 16286) -де ауыстырумен анықталған (Salas және басқалар, 2002), енді кодтау аймағының маркерімен (np 3918) қолдау табады (сурет 2A) және алтыдан төртеуінде табылған. Йемен L2a1 тегі. L2a1a Африканың оңтүстік-шығысында ең жоғары жиілікте кездеседі (Перейра және басқалар 2001; Салас және басқалар 2002). Олардың арасында негізін қалаушы гаплотип те, туындылар да бар (16092 мутациямен) Йемендіктер ішінде нақты сәйкестіктер бар Мозамбик реттіліктер (Перейра және басқалар 2001; Салас және басқалар 2002). L2a1a солтүстік-батыс Африкада аз жиілікте кездеседі Мауре және Бамбара туралы Мали және Мавритания.[20] (Рандо т.б. 1998; Мака-Мейер т.б. 2003)
L2a1a1 хаплогруппасы
L2a1a1 6152C, 15391T, 16368C маркерлерімен анықталады
L2a1b Haplogroup
L2a1b 16189 және 10143-тегі алмастырулармен анықталады. 16192 L2a1b және L2a1c-де жиі кездеседі; ол Египетте Солтүстік Африкада пайда болады, Африканың оңтүстік-шығысында да кездеседі, сондықтан ол сонымен бірге Бантудың кеңеюі.[5]
Haplogroup L2a1c
L2a1c көбінесе 16189 мутациясын L2a1b-мен бөліседі, бірақ 3010 және 6663-те өз маркерлері бар. 16192 L2a1b және L2a1c-де жиі кездеседі; ол Африканың оңтүстік-шығысында, сондай-ақ Африкада пайда болады.[21] Бұл осы кладусты in situ-де әртараптандыруды ұсынады.
L2a1 үстіндегі T16209C C16301T C16354T позициялары Кивисильд және басқалармен L2a1c деп аталатын кіші ішкі қабатты анықтайды. (2004, 3-сурет) (Salas және басқалардағы 6-суретті қараңыз), ол негізінен пайда болады Шығыс Африка (мысалы, Судан, Нубия, Эфиопия ) арасында Туркана және Батыс Африка (мысалы, Канури ).
Ішінде Чад бассейні, төрт түрлі L2a1c типтері Шығыс және Батыс Африка типтерінен бір немесе екі мутациялық қадамдармен анықталды. (Кивисилд және басқалар) 2004 ж.[21] (9 немесе 443 беттегі сілтеме)[22][23]
L2a1c1 Haplogroup тобы
L2a1c1 Африкадан шыққан.[24] Ол 198, 930, 3308, 8604, 16086 маркерлерімен анықталған. Тунисте Сефардтық, Ашкенази, еврейлер, еврейлер, марокколықтар, мысырлықтар, нубиялықтар және йемендіктерде байқалады.
Haplogroup L2a1f
Хосян, Замбия, Мадагаскар
L2a1k гаплогруппасы
L2a1k G6722A және T12903C маркерлерімен анықталады. Ол бұрын L2a1a еуропалық спецплед ретінде сипатталған және анықталған Чехтар және Словактар.[25]
L2a1l2a хаплогруппасы
L2a1l2a «ретінде танылдыАшкенази Орталық және Шығыс Еуропада ата-бабалары бар ашкенази еврейлерінің арасында кездесетін «гаплотоп. Сонымен қатар, бұл еврей емес поляк популяцияларында аз мөлшерде анықталды, ол Ашкенази қоспасынан шыққан деп болжануда.[26] Алайда, бұл гаплотип Ашкенази митохондрия шежіресінің өте аз үлесін ғана құрайды; әр түрлі зерттеулер (оның ішінде Бехар) оның жиілігін 1,4-1,6% аралығында көрсетті.
L2a2 Haplogroup
L2a2 үшін тән Мбути пигмийлері.[9]
Haplogroup L2b'c
L2b'c шамамен 62000 жыл бұрын дамыған.[1]
Haplogroup L2b
Бұл субклайд негізінен табылған Батыс Африка, бірақ ол бүкіл Африкада таралған.[27]
Haplogroup L2c
L2c көбінесе Батыс Африкада кездеседі және сол жерде пайда болуы мүмкін.[15] Арнайы бар Сенегал 39% -да, Кабо-Верде 16% және Гвинея-Бисау 16%.[6]
Haplogroup L2d
L2d көбінесе Батыс Африкада пайда болуы мүмкін.[15] Ол Йеменде, Мозамбикте және Суданда да кездеседі.[14]
Haplogroup L2e
L2e (бұрынғы L2d2) -ге тән Батыс Африка.[5] Ол сондай-ақ табылған Тунис,[28] және арасында Mandinka халқы бастап Гвинея-Бисау және афроамерикалықтар.[27]
Ағаш

L2 гаплогруппасының подкладтарының филогенетикалық ағашы Маннис ван Овен мен Манфред Кайсердің қағазына негізделген. Әлемдік митохондриялық ДНҚ вариациясының жан-жақты филогенетикалық ағашы жаңартылды[3] және кейінгі жарияланған зерттеулер.
- Ең соңғы ата-бабалар (MRCA)
- L1'2'3'4'5'6
- L2'3'4'6
- L2
- L2a'b'c'd
- L2a
- L2a1
- L2a1a
- L2a1a1
- L2a1a2
- L2a1a2a
- L2a1a2a1
- L2a1a2b
- L2a1a2a
- L2a1a3
- 16189 (16192)
- L2a1b
- L2a1b1
- L2a1f
- L2a1f1
- L2a1b
- 143
- L2a1c
- L2a1c1
- L2a1c2
- L2a1c3
- L2a1c4
- L2a1d
- L2a1e
- L2a1e1
- L2a1сағ
- 16189
- L2a1i
- L2a1j
- L2a1k
- 16192
- L2a1l
- L2a1l1
- L2a1l1a
- L2a1l2
- L2a1l1
- L2a1l
- L2a1c
- L2a1a
- L2a2
- L2a2a
- L2a2a1
- L2a2b
- L2a2b1
- L2a2a
- L2a1
- L2b'c
- L2b
- L2b1
- L2b1a
- L2b1a2
- L2b1a3
- L2b1a
- L2b1
- L2c
- L2c2
- L2c2a
- L2c3
- L2c2
- L2b
- L2d
- L2d1
- L2d1a
- L2d1
- L2a
- L2e
- L2a'b'c'd
- L2
- L2'3'4'6
- L1'2'3'4'5'6
Сондай-ақ қараңыз
| Wikimedia Commons-та бұқаралық ақпарат құралдары бар Haplogroup L2 (mtDNA). |
- Генеологиялық ДНҚ-тест
- Генетикалық шежіре
- Адамның митохондриялық генетикасы
- Популяция генетикасы
- Адамның митохондриялық ДНҚ гаплогруппалары
|
Филогенетикалық ағаш адамның митохондриялық ДНҚ-сы (mtDNA) гаплогруппалары | |||||||||||||||||||||||||||||||||||||||
| Митохондриялық Хауа (L ) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1-6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| М | N | ||||||||||||||||||||||||||||||||||||||
| CZ | Д. | E | G | Q | O | A | S | R | Мен | W | X | Y | |||||||||||||||||||||||||||
| C | З | B | F | R0 | алдын-ала JT | P | U | ||||||||||||||||||||||||||||||||
| HV | JT | Қ | |||||||||||||||||||||||||||||||||||||
| H | V | Дж | Т | ||||||||||||||||||||||||||||||||||||
Әдебиеттер тізімі
- ^ а б c г. Соареш, Педро; Лука Эрмини; Ноэль Томсон; Мару Мормина; Тереза Рито; Арне Рюл; Антонио Салас; Стивен Оппенгеймер; Винсент Маколей; Мартин Б. Ричардс (4 маусым 2009). «Селекцияны тазарту үшін түзету: жақсартылған адамның митохондриялық молекулалық сағаты». Американдық генетика журналы. 84 (6): 82–93. дои:10.1016 / j.ajhg.2009.05.001. PMC 2694979. PMID 19500773. Алынған 2009-08-13.
- ^ Силва, Марина (2015). «Орталық Африка митохондриялық гаплогруппасы L2 тіркелген Орталық және Шығыс Африка арасындағы 60 000 жылдық өзара іс-қимыл». Ғылыми баяндамалар. Табиғат. 5: 12526. Бибкод:2015 Натрия ... 512526S. дои:10.1038 / srep12526. PMC 4515592. PMID 26211407.
- ^ а б ван пеші, Маннис; Манфред Кайсер (2008 ж. 13 қазан). «ДНҚ-ның ғаламдық вариациясының жан-жақты филогенетикалық ағашы». Адам мутациясы. 30 (2): E386-E394. дои:10.1002 / humu.20921. PMID 18853457. S2CID 27566749. Архивтелген түпнұсқа 2012 жылғы 4 желтоқсанда. Алынған 2009-05-20.
- ^ Тишкофф және басқалар, бүкіл-mtDNA геномдық дәйектіліктің антикалық африкалық шежіресін талдау, Молекулалық биология және эволюция, т. 24, жоқ. 3 (2007), 757-768 б.
- ^ а б c г. e f Салас, Антонио және басқалар. Африка mtDNA ландшафтының жасалуы, Американдық генетика журналы, т. 71, жоқ. 5 (2002), 1082–1111 бб.
- ^ а б c Роза, Александра; Брем, А; Кивисильд, Т; Metspalu, E; Виллемс, Р; т.б. (2004). «Батыс Африка Гвинеяларының MtDNA профилі: Сенегамбия аймағын жақсы түсінуге». Адам генетикасының жылнамалары. 68 (Pt 4): 340-352. дои:10.1046 / j.1529-8817.2004.00100.x. PMID 15225159. S2CID 15391342.
- ^ Адам саяхаты атласы: Haplogroup L2 Мұрағатталды 2011-10-06 сағ Wayback Machine Генографиялық жоба, National Geographic.
- ^ Силва, Марина; Алшамали, Фарида; Сильва, Паула; Каррилью, Карла; Мандат, Флавио; Иса Тровоада, Мария; Жерный, Виктор; Перейра, Луиса; Соареш, Педро (2015). «Орталық Африка митохондриялық гаплогруппасы L2 тіркелген Орталық және Шығыс Африка арасындағы 60 000 жылдық өзара іс-қимыл». Ғылыми баяндамалар. 5: 12526. Бибкод:2015 Натрия ... 512526S. дои:10.1038 / srep12526. PMC 4515592. PMID 26211407.
- ^ а б Кинтана-Мурчи және басқалар. 2008 ж. Пигмийлік аңшылар мен банту тілінде сөйлейтін фермерлер арасында терең ата-баба мен асимметриялық гендердің аналық іздері. 'Америка Құрама Штаттарының Ұлттық ғылым академиясының еңбектері'. 105 (5): 1599
- ^ а б Сади Андерсон-Манн 2006, Африка митохондриялық ДНҚ вариациясының филогенетикалық және филогеографиялық анализі. Мұрағатталды 2011-09-10 сағ Wayback Machine
- ^ а б Veeramah, Krishna R және басқалар 2010, Нигерияның Кросс-өзен аймағындағы халықтарда тілдің едәуір өзгеруі болған кезде, бірпарентерлік маркерлермен бағаланған генетикалық дифференциация.
- ^ Сирак, Кендра; Френандес, Даниэль; Новак, Марио; Ван Гервен, Деннис; Пинхаси, Рон (2016). «IUAES 2016 конгресс аралық реферат кітабы - бөлінген қауым? Ортағасырлық Кулубнарти қоғамдастық геномын (-тарын) келесі буын тізбегін қолдану арқылы ашу». Iuaes конгрессаралық реферат кітабы 2016 ж. IUAES: 115.
- ^ Серезо, Мария; т.б. (2011). «Чад көлінің бассейнінің популяциялық құрылымы туралы жаңа түсініктер SNP кодтайтын митохондриялық ДНҚ-ны генотиптеу арқылы анықталды». PLOS ONE. 6 (4): e18682. Бибкод:2011PLoSO ... 618682C. CiteSeerX 10.1.1.291.8871. дои:10.1371 / journal.pone.0018682. PMC 3080428. PMID 21533064.
- ^ а б c Тоомас Кивисилд және басқалар, Эфиопиялық Митохондрияның ДНҚ мұрасы: Ген ағынын Көз жасының қақпасы бойынша және айналасында қадағалау, Американдық генетика журналы, т. 75, жоқ. 5 (2004 ж. Қараша), 752–770 бб.
- ^ а б c г. Антонио Торрони және басқалар, mtDNA Haplogroup L2 төрт қабаты әр түрлі деңгейде дамиды ма ?, Американдық генетика журналы, т. 69 (2001), 348-1356 бб.
- ^ Кампира, Е; Кумвенда, Дж; ван Оостерхут, Джейдж; Дандара, С (тамыз 2013). «Митохондриялық ДНҚ субгаплогруппалары L0a2 және L2a малавиялық ересектердегі перифериялық нейропатияға сезімталдығын ставудин құрамында белсенділігі жоғары антиретровирустық терапия бар». J иммундық тапшылық синдромын сатып алыңыз. 63 (5): 647–652. дои:10.1097 / QAI.0b013e3182968ea5. PMC 3815091. PMID 23614993.
- ^ Жоқ, Эми. «ГЕНЕТИКАЛЫҚ МӘЛІМЕТТЕРДІ ЖАҢА АДАМДЫҢ ЭВОЛЮЦИЯЛЫҚ ТАРИХЫ ЖӘНЕ КҮЙІНДІ АУРУЫН ТЕРГЕУ ҮШІН ПӘН БОЙЫНША ШЕШТІҢ ТАЛДАУЫ» (PDF). Флорида университеті. Алынған 2 қараша 2016.
- ^ Манко, Жан (2013). Ата-бабалардың саяхаттары: Еуропаның алғашқы венерлерінен Викингтерге дейінгі популярлық кезеңі. Темза және Хадсон. б. 88. ISBN 978-0500771822. Алынған 29 қыркүйек 2017.
- ^ Скоглунд; т.б. (2017 жылғы 21 қыркүйек). «Тарихқа дейінгі африкалық халықтың құрылымын қалпына келтіру». Ұяшық. 171 (1): 59–71. дои:10.1016 / j.cell.2017.08.049. PMC 5679310. PMID 28938123. Алынған 15 қазан 2017.
- ^ González, A. M. және басқалар, 2006, Мавритания мен Малидегі митохондриялық ДНҚ-ның өзгеруі және олардың басқа Батыс Африка популяцияларымен генетикалық байланысы
- ^ а б «Wiley Онлайн Кітапханасы | Ғылыми зерттеу мақалалары, журналдар, кітаптар және анықтамалық жұмыстар». Архивтелген түпнұсқа 2010-08-05. Алынған 2009-05-19.
- ^ Керни, V және басқалар 2006, Сахел-Судан белдеуіндегі екі бағытты дәліз және Чад бассейні популяцияларының айрықша ерекшеліктері: Митохондриялық ДНҚ геномы ашқан тарих.
- ^ Ласкаро, Даниэла; кастельхана, Стефано; Гаспарре, Джузеппе; Ромео, Джованни; Сакконе, Сесилия; Attimonelli, Marcella (2008). «RHNumtS компиляциясы: Адам NumtS-ті анықтау мен сандық анықтауға арналған биоинформатика ерекшеліктері мен тәсілдері». BMC Genomics. 9: 267. дои:10.1186/1471-2164-9-267. PMC 2447851. PMID 18522722. Алынған 12 сәуір 2020.
- ^ Ласкаро, Даниэла; Кастеллана, Стефано; Гаспарре, Джузеппе; Ромео, Джованни; Сакконе, Сесилия; Attimonelli, Marcella (2008). «RHNumtS компиляциясы: Адам NumtS-ті орналастыру және олардың мөлшерін анықтауға арналған биоинформатика ерекшеліктері мен тәсілдері». BMC Genomics. 9: 267. дои:10.1186/1471-2164-9-267. PMC 2447851. PMID 18522722.
- ^ Борис А Малярчук, Мирослава Деренко, Мария Перкова, Томаш Гзыбовски, Томас Ванекек және Ян Лазур, славяндарда африкалық митохондриялық ДНҚ тектес филогениясын қалпына келтіру, Еуропалық адам генетикасы журналы, т. 16 (2008), 1091–1096 бб
- ^ Марта Мельник-Сикорска, Патрича Дака, Борис Малярчук, Мирослава Деренко, Катарзина Скониечна, Мария Перкова, Тадеуш Добош, Томаш Гзыбовски, Митохондриялық геномның толық тізбегінен алынған славяндар тарихы PLOSO 2013 жылдың 14 қаңтарында; 10.1371 / journal.pone.0054360
- ^ а б Бехар және басқалар 2008b, Адамның матрилиалдық алуан түрлілігінің таңы Am J Hum Genet. 2008 жылғы 9 мамыр; 82 (5): 1130–1140
- ^ Коста MD және басқалар, 2009 Тунис жүзжылдықтарының толық mtDNA тізбегінен алынған деректер: гаплогруппаның қауымдастығы және ұзақ өмірге «алтын орта». ([1] )
Сыртқы сілтемелер
- Ян Логандікі Haplogroup L2. Митохондриялық ДНҚ орны
- Ян Логандікі L2bcd Митохондриялық ДНҚ орны
- Маннис ван пеші PhyloTree.org - mtDNA кіші ағашы L
- H2 блогының таралуы L2, бастап ұлттық географиялық