Циклинге тәуелді киназа кешені - Cyclin-dependent kinase complex
A циклинге тәуелді киназа кешені (CDKC, циклин-CDK) Бұл ақуыз кешені ақуыз киназының белсенді емес каталитикалық суббірлігі ассоциациясынан пайда болған, циклинге тәуелді киназа (CDK), нормативті бөлімшемен, циклин.[1] Циклинге тәуелді киназалар циклинмен байланысқаннан кейін түзілген кешен активтендірілген күйде болады. Активтендірілген кешеннің субстрат ерекшелігін негізінен кешен ішіндегі ассоциацияланған циклин анықтайды. CDKC белсенділігі мақсатты ақуыздардың фосфорлануымен, сондай-ақ ингибиторлық белоктардың байланысуымен бақыланады.[2]
Құрылымы мен ережелері

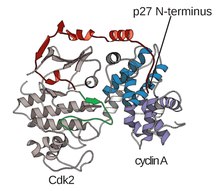
Құрылымы CDK Циклинді суббірліктермен (CDKC) кешенде құрылымдық және жасушалық биологтардың мақсаты 1990 жылдан бастап байланыспаған А циклинінің құрылымын Браун және басқалар шешкен кезден басталды. және сол жылы Джефери және басқалар. адамның циклині A-CDK2 кешенінің құрылымын 2,3 Ангстромға дейін шешті.[3] Осы уақыттан бастап көптеген CDK құрылымдары жоғары ажыратымдылыққа ие болды, олардың ішінде әр түрлі субстраттармен байланысқан CDK2 және CDK2 құрылымдары бар, жоғары ажыратымдылық құрылымдары шамамен 25 CDK-циклинді кешендер үшін барлығы бар Ақуыздар туралы мәліметтер банкі.[4] Функция негізінде CDK-циклинді кешенді құрылымдардың екі жалпы популяциясы бар, олар ашық және жабық формада. Пішіндер арасындағы айырмашылық циклиндік серіктестердің байланысында болады, мұнда тұйық формалы кешендер CDK-циклинді байланыстыру кезінде ЦДК активация циклінің C және N-терминалдарында байланысады, ал ашық форма серіктестері тек N-терминалда байланысады. Ашық пішінді құрылымдар көбінесе транскрипциялық реттеуге қатысатын кешендерге сәйкес келеді (CDK 8, 9, 12, 13), ал жабық түрдегі CDK-циклин кешені көбінесе жасуша циклінің прогрессиясы мен реттелуіне қатысады (CDK 1, 2, 6) . Бұл ерекше рөлдер, алайда, CDK компоненттері арасындағы реттілік гомологиясымен айтарлықтай ерекшеленбейді. Атап айтқанда, осы белгілі құрылымдардың арасында төрт негізгі консервіленген аймақ бар көрінеді: N-терминалы глицинге бай цикл, топса аймағы, αC-спираль және T-циклды реттеу орны.[4]
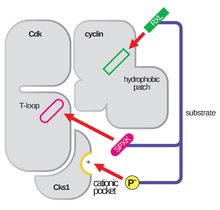
Іске қосу циклі
The белсендіру циклі, сонымен қатар T-цикл деп аталады, бұл CDK аймағы (көптеген CDK-да DFG және APE мотивтері арасында)[4] CDK функционалды серіктесімен байланысқан кезде ферментативті белсенді болады. CDK-циклин кешендерінде бұл активтену аймағы консервленген αL-12 спиралдан тұрады және құрамында фосфорланатын негізгі қалдық бар (әдетте Треонин CDK-циклинді серіктестер үшін, сонымен қатар, Серин мен Тирозин), CDK-дың ферментативті белсенділігімен айналысады. Дәл осы маңызды қалдықта (CDK2 кешендеріндегі T160, CDK6 комплекстеріндегі T177) CDK-циклинді комплекстердің ферментативті ATP-фосфорлануы CAK (циклинді белсендіретін киназа, адам жасушаларында CDK7-Циклин Н комплексіне сілтеме жасай отырып) жүреді. Осы учаскеде АТФ-ті фосфорилатқа дейін гидролиздегеннен кейін, бұл кешендер өз функцияларын, яғни ұялы нысандардың фосфорлануын аяқтай алады. CDK 1, 2 және 6-да T-цикл және жеке C-терминал аймағы ЦДК-да циклинмен байланысатын негізгі орындар болып табылатындығын және осы ЦДК-дің әрқайсысымен байланысқан циклиндер болып табылатындығын ескеру маңызды. T-циклін іске қосу торабының нақты бірізділігі. Бұл циклинді байланыстыратын учаскелер осы құрылымдық компоненттің αL-12 Helix мотивін қоршап тұрған салыстырмалы түрде жоғары гомологиясына қарамастан, CDK-дегі ең жоғары өзгергіштік аймақ болып табылады.[4]
Глицинге бай аймақ
The глицин CDK2-дегі 12-16 қалдықтарында байқалатын бай цикл (Gly-бай цикл) ашытқы түрінде де, жануарлар моделінде де сақталған GXGXXG мотивін кодтайды. Реттелетін аймақ осы мотив шеңберінде глицин емес қалдықтарда дифференциалды фосфорлануға ұшырайды, сондықтан бұл сайт осы объектіге бағынады У1 және / немесе Мыт1 ингибиторлық киназа фосфорлануы және Ccc25 сүтқоректілерде де-фосфорлану. CDK2-дегі Gly-бай циклдегі бұл қайтымды фосфорлану белсенділік одан әрі зерттелген Y15-те жүреді. Осы қалдықты зерттеу фосфорлану АДФ пен субстраттың КДК-циклинді кешендердің активация цикліндегі осы қажетті байланыстыру орындарымен стерикалық араласу арқылы байланысуына жол бермейтін конформациялық өзгеріске ықпал ететіндігін көрсетті. Бұл белсенділікке GL-ге бай циклдің CDK-циклин кешені құрылымының жалпы өзгеріссіз субстраттың жақындығын төмендетуге едәуір әсер ету үшін оның белсенділену цикліне қарай бұрылуына мүмкіндік беретін көптеген CDK құрылымындағы иілгіштік көмектеседі.[3][5]
Топса аймағы
Эукариоттық жасушалардың ішіндегі CDK-нің сақталған топса аймағы Gly-бай цикл мен активация циклы арасындағы маңызды көпір рөлін атқарады. CDK N-терминалды лобымен сипатталады, ол бірінші кезекте осы ілгек аймағы арқылы альфа-спираль үстемдігі бар C-терминалды лобқа қосылған бета-парағымен бұралған. Т-цикл және гли-бай циклды талқылай отырып, олардың биохимиялық функцияларын жүзеге асыру үшін кеңістіктік өзара әрекеттесуге қабілетті болуы керек бұл аймақтар КДК-нің қарама-қарсы қабаттарында жатқанын ескеру маңызды. Осылайша, CDK типі мен CDK-циклин кешені арасында ұзындығы аздап өзгеруі мүмкін бұл топса аймағы, осы үлестерді қосу арқылы CDK-нің маңызды реттеуші аймақтарын байланыстырады және ATP-ді дұрыс бағдарлай отырып, CDK-циклина кешендерінің алынған құрылымында негізгі рөлдерді атқарады. құрастырылған кешен арқылы фосфорлану реакцияларының жеңіл катализі үшін. [3][4]
αC-спираль
ΑC-Helix аймағы көптеген сүтқоректілер киномында (тұқымдасы) жоғары сақталған киназалар ). Оның негізгі жауапкершілігі - сақтау аллостериялық бақылау киназа белсенді учаскесінің. Бұл бақылау CDK-циклинді кешендерде CDK белсенділігін оның серіктес реттеушімен байланысқанға дейін (мысалы, циклин немесе басқа серіктес ақуыз) белсенділіктің алдын-алу арқылы көрінеді. Бұл байланыс CDK-нің αC-Helix аймағында конформациялық өзгерісті тудырады және оны белсенді учаске саңылауынан жылжытуға мүмкіндік береді және T-циклын іске қосудың бастапқы процесін аяқтайды. Бұл аймақ киназдардың суперфамилиясында соншалықты сақталғанын ескере отырып, αC-Helix киназаның N-терминалды лобынан бүктеліп, αL-12 спиралына қол жетімділікті арттыруға мүмкіндік беретін механизм көрсетілген. T-циклі есірткіні дамытудың әлеуетті мақсаты болып саналады.[6]
Жасушалық цикл
Ашытқы жасушаларының циклі
Бұл кешендер әр түрлі функцияларға ие болғанымен, CDKC-лер олардың рөлдерімен ең танымал жасушалық цикл. Бастапқыда зерттеулер жүргізілді Шизосахаромицес помбы және Saccharomyces cerevisiae (ашытқы) S. pombe және S. cerevisiae бір-біріне ұқсамайтын Cdk, Cdc2 және Cdc28 сәйкес келеді, олар бірнеше түрлі циклиндермен біріктіріледі.[7] Циклинге байланысты жасуша циклінің әр түрлі бөліктері әсер етеді. Мысалы, in S. pombe, Cdc2 Cdk13-пен байланысып, Cdk13-Cdc2 кешенін құрайды. Жылы S. cerevisiae, Cdc28 циклиндермен, Cln1, Cln2 немесе Cln3 ассоциациясы G1 фазасынан ауысуға әкеледі S фазасы. S фазасында бір рет Cln1 және Cln2 Cdc28-мен диссоциацияланып, Cdc28 мен Clb5 немесе Clb6 арасындағы комплекстер түзіледі. G2 фазасында Cdc28 мен Clb1, Clb2, Clb3 немесе Clb4 арасындағы ассоциациядан түзілген комплекстер, прогрессияға әкеледі G2 фаза M (митоздық) фазаға дейін. Бұл кешендер ерте М фазасында да бар.[2] Ашытқы CDKC-нің қысқаша мазмұнын 1-кестеден қараңыз.
- Кесте 1. Ашытқыдағы жасуша циклінің фазаларымен байланысты CDKC
| CDK | Циклин | Ұяшық циклінің фазасы |
|---|---|---|
| CD2 (S. pombe) | Cdc13 | G2-ден M фазасына өту; ерте М фазасы |
| CD28 (S. cerevisiae) | Cln1, Cln2 | G1-ден S фазалық ауысу |
| Cdc28 | Clb5, Clb6 | S фазасы |
| Cdc28 | Clb1, Clb2, Clb3, Clb4 | G2-ден M фазасына өту; ерте М фазасы |
Ашытқыдағы жасуша циклінің әр фазасында пайда болған кешендер туралы белгілі болған маңызды фосфорлану орындары мен транскрипция факторларына негізделген ұсынылған модельдер пайда болды.[7][8]
Сүтқоректілердің жасушалық циклі
Ашытқы жасушаларының циклін зерттеу нәтижесінде анықталған ақпаратты қолдану арқылы сүтқоректілердің жасушалық циклына қатысты айтарлықтай жетістіктерге қол жеткізілді. Жасуша циклдарының ұқсас екендігі анықталды және CDKC тікелей немесе жанама түрде жасуша циклінің прогрессиясына әсер етеді. Бұрын айтылғандай, ашытқыларда тек бір циклинге тәуелді киназа (CDK) бірнеше түрлі циклиндермен байланысады. Алайда, сүтқоректілердің жасушаларында бірнеше түрлі ЦДК әр түрлі циклиндермен байланысып, CDKC түзеді. Мысалы, Cdk1 (адам Cdc2 деп те аталады), адамның алғашқы CDK анықталған циклиндермен байланысады A немесе B. CyclinA / B-Cdk1 кешендері G2 фазасы мен M фазасы, сондай-ақ ерте M фазасы арасындағы ауысуды басқарады. Сүтқоректілердің тағы бір CDK, Cdk2, D1, D2, D3, E немесе A. циклиндерімен кешендер түзе алады. Cdk4 және Cdk6 D1, D2 және D3 циклиндерімен әрекеттеседі.[9] Зерттеулер көрсеткендей, CDKC циклінің D1-Cdk4 / 6 арасында ешқандай айырмашылық жоқ, сондықтан кез-келген ерекше қасиеттер субстраттың ерекшелігімен немесе активациясымен байланысты болуы мүмкін.[1] CDK деңгейлері жасушаның бүкіл циклінде біршама тұрақты болып тұрса, циклин деңгейлері өзгеріп отырады. Флуктуация циклин-CDK кешендерінің активтенуін және ақыр соңында бүкіл циклде прогрессияны басқарады.[10] Жасушалық циклге қатысқан сүтқоректілердің CDKC жасушаларының қысқаша мазмұнын 2-кестеден қараңыз.
- Кесте 2. Сүтқоректілер жасушасындағы жасушалық цикл фазасымен байланысты CDKC[4]
| CDK | Циклин | Ұяшық циклінің фазасы | Циклинді емес серіктес ақуыздар |
|---|---|---|---|
| Cdk1 (Cdc2) | А және В циклиндері | G2-ден M фазасына өту; ерте М фазасы | Cks1 және Cks2 |
| Cdk2 | D1, D2, D3 циклиндері | G1 фазасы | KAP, Cks1, p27KIP1 және Spy-1 |
| Cdk2 | Циклин Е | G1-ден S фазалық ауысу | KAP, Cks1, p27KIP1 және Spy-1 |
| Cdk2 | Циклин А | S фазасы | KAP, Cks1, p27KIP1 және Spy-1 |
| Cdk4 | D1, D2, D3 циклиндері | G1 фазасы | HSP90-Cdc37 |
| Cdk6 | D1, D2, D3 циклиндері | G1 фазасы | p16INK4A, p19INK4D және P18INK4C-циклин K |
| Cdk8 | Cyclin C | --- | --- |
| CD9 | Циклин Т | --- | Tat, AFF4 және TAR |
| Cdk12 | Циклина К | --- | --- |
| Cdk13 | Циклина К | --- | --- |

G1 S фазалық прогрессияға дейін
Соңында Г.1 фазасы, CDKCs байланыстырады және фосфорилат мүшелерін ретинобластома (Rb) ақуызы отбасы. Rb ақуыздар тобының мүшелері - ісіктердің супрессорлары, олар ісік пайда болған кезде пайда болатын жасушалардың бақыланбайтын көбеюін болдырмайды. Сонымен қатар, pRbs G-дан ауысу үшін қажетті гендерді басады деп ойлайды1 фазадан S фазаға дейін. Жасуша келесі фазаға өтуге дайын болған кезде CDKCs, D1-Cdk4 циклині және D1-Cdk6 фосфорилат pRB циклиндері, одан кейін E-Cdk2 CDKC циклинінен қосымша фосфорлану жүреді.[11][12] Фосфорлану пайда болғаннан кейін, транскрипция факторлары содан кейін рРБ-ны қайтымсыз инактивациялау үшін шығарылады және жасуша циклінің S фазасына прогрессия басталады.[13] G-да қалыптасқан E-Cdk2 CDKC циклині1 фаза содан кейін S фазасы кезінде ДНҚ репликациясын бастауға көмектеседі.[1]
G2 М фазасының прогрессиясына дейін
S фазасының соңында А циклині Cdk1 және Cdk2-мен байланысады. G2 фазасы кезінде А циклині ыдырайды, ал B циклині синтезделіп, B-Cdk1 циклиндері түзіледі. М фазасына өту үшін B-Cdk1 циклин кешендері маңызды болып қана қоймай, бұл CDKC келесі реттеуші және құрылымдық процестерде маңызды рөл атқарады:[1]
- Хромосомалық конденсация
- Гольджи желісінің фрагментациясы
- Ядролық ламинаның бұзылуы
B циклинінің деградациясы арқылы B-Cdk1 кешенін инактивациялау жасуша циклінің M фазасынан шығу үшін қажет.[1]
Басқа
Белгілі CDKC-дің көп бөлігі жасуша циклына қатысса да, барлық киназа кешендері осылай жұмыс істемейді. Зерттеулер көрсеткендей, циклин k-Cdk9 және T1-Cdk9 циклині сияқты басқа CDKC-лер қатысады. репликациялық стресс жауап,[14] және ықпал ету транскрипция.[15][16][17] Сонымен қатар, H-Cdk7 циклин кешендері де рөл атқаруы мүмкін мейоз аталық жыныс жасушаларында,[18] және транскрипциялық іс-шараларға қатысатындығы көрсетілген.[1][19]
Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ а б c г. e f Malumbres M, Barbacid M. Сүтқоректілердің циклинге тәуелді киназалары. Трендтер биохимия. Ғылыми. 2005 қараша; 30 (11): 630-41
- ^ а б Лодиш Х, Балтимор Д, Берк А, Зипурский С.Л., Мацудайра П, Дарнелл Дж. 1995. Молекулалық жасуша биологиясы. 3-ші басылым. Нью-Йорк: Американдық ғылыми кітаптар
- ^ а б c Кристи Левин, Фредерик Р. Кросс, құрылымдық жасушалық цикл биологиясы, құрылым, 3 том, 11-шығарылым, 1995 ж., 1131-1134 беттер, ISSN 0969-2126, https://doi.org/10.1016/S0969-2126(01)00248 -9.
- ^ а б c г. e f Wood, D. J., & Endicott, J. A. (2018). CDK-циклиндер отбасының функционалды әртүрлілігі туралы құрылымдық түсініктер. Ашық биология, 8 (9), 180112.
- ^ Малумбралар: циклинге тәуелді киназалар. Геном биологиясы, 2014, 15:22, доии: 10.1186 / gb4184
- ^ Лоренцо Палмиери, Джулио Растелли, спиральдың αC жылжуы ақуыз киназдарының аллостериялық модуляциясының жалпы тәсілі ретінде, есірткінің табылуы Бүгін, 18 том, 7-8 шығарылым, 2013, 407-414 беттер, ISSN 1359-6446, https: // doi. org / 10.1016 / j.drudis.2012.11.009.
- ^ а б Саймон I, Барнетт Дж, Ханнетт Н, Харбисон КТ, Риналди Н.Ж., Волкерт Т.Л., Вайрик Дж.Дж., Цейтлингер Дж, Гиффорд Д.К., Яаккола Т.С., Янг РА. Ашытқы жасушаларының цикліндегі транскрипциялық реттегіштердің сериялық реттелуі. Ұяшық. 2001 қыркүйек 21; 106 (6): 697-708.
- ^ Барик Д, Бауманн ВТ, Пол М.Р., Новак Б, Тайсон Джейдж. Мультиситті фосфорлануға негізделген ашытқы жасушаларының циклін реттеу моделі. Mol Syst Biol. 2010 ж. 24 тамыз; 6: 405.
- ^ Malumbres M, Barbacid M. Жасуша циклі, CDK және қатерлі ісік: өзгеретін парадигма. Nat Rev Cancer. 2009 наурыз; 9 (3): 153-66.
- ^ Вермюлен К, Ван Бокстаеле Д.Р., Бернеман З.Н. Жасушалық цикл: қатерлі ісік кезіндегі реттелуді, реттелуді және терапевтік мақсатты шолу. Жасуша Пролиф. 2003 маусым; 36 (3): 131-49.
- ^ Mittnacht S. pRB фосфорлануын бақылау. Curr Opin Genet Dev. 1998 ақпан; 8 (1): 21-7.
- ^ Каелин ВГ кіші ретинобластома ақуызының қызметтері. Биосессиялар. 1999 қараша; 21 (11): 950-8.
- ^ Лундберг А.С., Вайнберг Р.А. Ретинобластома ақуызының функционалды инактивациясы кем дегенде екі бөлек циклин-cdk кешенінің дәйекті түрленуін қажет етеді. Mol Cell Biol. 1998 ақпан; 18 (2): 753-61.
- ^ Yu DS, Zhao R, Hsu EL, Cayer J, Ye F, Guo Y, Shyr Y, Cortez D. Циклинге тәуелді киназа 9-циклин K репликация стрессінде жауап береді. EMBO Rep. 2010 қараша; 11 (11): 876-82.
- ^ Fu TJ, Peng J, Lee G, Price DH, Flores O. Cyclin K CDK9 реттеуші суббірлігі ретінде жұмыс істейді және РНҚ полимераза II транскрипциясына қатысады. J Biol Chem. 1999 3 желтоқсан; 274 (49): 34527-30.
- ^ Ян З, Чжу Q, Луо К, Чжоу Q. 7SK кіші ядролық РНҚ транскрипцияны бақылау үшін CDK9 / циклин T1 киназасын тежейді. Табиғат. 2001 15 қараша; 414 (6861): 317-22.
- ^ Ю DS, Cortez D. CDK9-циклин К геномының тұтастығын сақтаудағы рөлі. Ұяшық циклі. 2011 1 қаңтар; 10 (1): 28-32.
- ^ Kim JM, McGaughy JT, Bogle RK, Ravnik SE. Тінтуірдің аталық жыныс жасушаларында циклин H / Cdk7 кешенінің мейоздық экспрессиясы. Biol Reprod. 2001 мамыр; 64 (5): 1400-8.
- ^ Patel SA, Simon MC. Тышқан эмбриональды дің жасушалары мен эмбриондарындағы Cdk7.cyclin H.Mat1 кешенін функционалды талдау. J Biol Chem. 2010 мамыр 14; 285 (20): 15587-98.