Тауан бүркіт - Tawny eagle
| Тауан бүркіт | |
|---|---|
 | |
| Қайдан Этоша ұлттық паркі | |
| Ғылыми классификация | |
| Корольдігі: | Анималия |
| Филум: | Chordata |
| Сынып: | Aves |
| Тапсырыс: | Accipitriformes |
| Отбасы: | Accipitridae |
| Тұқым: | Акила |
| Түрлер: | A. rapax |
| Биномдық атау | |
| Акила рапаксы (Темминк, 1828) | |
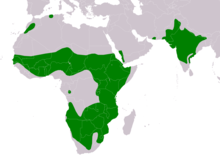 | |
| Диапазоны A. rapax Тұрғын | |
| Синонимдер | |
Акила рапаксы | |

The жалаңаш бүркіт (Акила рапаксы) үлкен, ұзақ өмір сүреді жыртқыш құс. Барлығы сияқты бүркіттер, бұл отбасына жатады Accipitridae. Оның қатты қауырсынды аяқтары оны субфамилияның мүшесі ретінде көрсетеді Аквилиналар, «бүркіт» деп те аталады.[2] Тауан бүркіттері көбінесе олардың ауқымын құрайтын кең, бірақ үзілісті тұқымға ие Африка континент, сондай-ақ Үнді субконтиненті, сирек резиденттік оңтүстігінде орын алады Таяу Шығыс.[3] Оның ішінде ауқымы, бұл ашық құрғақтықты қолдайды тіршілік ету ортасы сияқты жартылай серуендер, шөлдер дала, немесе саванна жазықтар. Бағыттарына артықшылық бергеніне қарамастан құрғақшылық, түр сирек кездеседі, олар ағаштар мүлдем жоқ жерлерде.[4] Бұл бір-үштен тұратын резидент селекционер жұмыртқа ішінде таяқ ұя көбінесе ағаштың тәжінде болады.[5] Жалаңаш бүркіт оның барлық таксономиялық кландарының ішіндегі ең жоғары оппортунисті болып табылады және оны жиі аулайды өлексе немесе айналысады клептопаразитизм басқа жыртқыш жануарларға қатысты, сонымен бірге батыл және белсенді жыртқыш, көбінесе салыстырмалы түрде үлкен және алуан түрлі жыртқыш.[6] Есептеулерге қарағанда, бүркіттер 16 жасқа жетуі мүмкін.[7] Алайда, бүркіттің барлық таралу аймағында күрт төмендеу байқалды. Көптеген факторлар, әсіресе ұя салудың жоғалуы ағаш кесу және ғаламдық жылуы, Сонымен қатар қудалау (негізінен улану ) және басқа да антропогендік өлім (көбіне әртүрлі қолдан жасалған заттармен жанасу арқылы) бір кездегі көптеген саяхатшы бүркітті құрып кетудің аз-ақ алдында шығарады.[1][7][8]
Таксономия
Голландия натуралисті Коенраад Джейкоб Темминк суретті бүркітті 1828 жылы үндістаннан суреттеді үлгі үлгісі.[9] «Таван бүркіт» ресми атауы болып тағайындалды Халықаралық орнитологтар одағы (ХОК).[10] Акила болып табылады Латын ал «бүркіт» үшін рапакс латын тілінен аударғанда «ұғу» деген мағынаны білдіреді және рапас сөзіне қатысты, яғни өте жыртқыш.[5][11][12] Тауық бүркіт етік бүркіт ішіндегі кіші отбасы Accipitridae отбасы.[3] Жүктелген бүркіт кланы монофилетикалық және зерттеу кариотиптер олардың жалпы экскиппидридтер отбасында жақын және аз сыртқы байланыстары болуы мүмкін екенін көрсетті.[2][13][14] Бүркіттің субфамилиясының барлығының аяқтарын жауып тұратын қауырсындары бар және олар аккипитридтерден тұратын барлық континенттерде таралған.[6] Тұқым Акила дәстүрлі түрде ашық, ашық, қараңғы және ұзын қанатты бүркіттер ретінде анықталды.[15] Алайда, зерттеу генетика бірнеше қайта жіктеуге әкелді. Оларға кішкентай, бозарған және орманда мекендейтін бүркіттердің жылжуы жатады Акила, бірегейдің бөлінуі дала бүркіттері тұқымға Кланга сонымен қатар кейбір кішкентай бүркіттердің тұқымға бөлінуі Hieraeetus тыс Акила. Сонымен қатар, генетикалық зерттеулер жүргізілген бүркіт пен оның жақын туыстарының арасындағы үстірт ұқсас бүркіттердің және басқа да үстірт ұқсастықтың араздығын анықтады. Акила сияқты бүркіт (Aquila chrysaetos) түрлік кешен. Осы уақытқа дейін бүркіт түрлерінің тобы және алтын бүркіттердің тобы жақын туыстықтың жоқтығына қарамастан, сол тұқымдаста сақталады.[2][15][16][17]

Жалаңаш бүркіт бұған дейін қарастырылған ерекше бірге көші-қон дала бүркіті (Aquila nipalensis).[6][18] Бұл бүркіттер 1991 жылдың өзінде-ақ бір түрдің бір бөлігі болып саналды.[19] Морфология мен анатомиядағы айқын айырмашылықтар негізінде дала мен аңшы бүркіттер екіге бөлінді. Дала бүркіті - әлдеқайда айқынырақ, үлкенірек құс тегістеу, және екі бүркіттің бір-біріне түсініксіз ұқсастығына қарамастан, дамудың барлық кезеңдерінде түрлі-түсті болуымен, өзінің қанатты жақтауларымен, үлкен қанаттарымен және дәлелдерімен ерекшеленеді.[20][21][22][23][24] Сонымен қатар, тиісті түрлер экологиясы бойынша ерекшеленеді (диеталық биология, ұя салу әдеттері) және қатты аллопатикалық олардың асыл тұқымды жерлерінің таралуында.[5][20] Екі молекулалық зерттеу, олардың әрқайсысы гендердің өте аз мөлшеріне негізделген, түрлердің бір-бірінен айырмашылығы бар, бірақ олардың қаншалықты тығыз байланыста екендігі туралы келіспеушіліктер бар.[25][26] Генетикалық тұрғыдан алғанда, жалаңаш бүркіт жақынырақ шоғырлануы мүмкін империялық бүркіт түрлік кешен дала бүркіті көп болғанымен симпатикалық солтүстік еуразиялық бүркіттермен.[2]
Түршелер

Таңғы бүркіттердің үш сипаттамасы бар. Түрлердің түршелік жіктелуі кейде әртүрлі морфтардың вариациялары мен тіршілік етуімен күрделі болып саналды, өз кезегінде олар бір кездері шартты болып саналды.[3] Алайда, әр кіші түр географияда негізінен аллопатиялық болып табылады, тек солтүстік бөлігінде орналасқан негізгі түсініксіздіктер Африканың шығысы мұнда екі африкалық нәсіл де деңгейге көтерілуі мүмкін.[27]
- A. r. рапакс; Африкада оңтүстіктен таралған Конго Демократиялық Республикасы және орталық Кения оңтүстікке қарай.[3] Алайда бұл жарыс солтүстікке дейін созылуы мүмкін Эфиопия сонымен қатар (осылайша Эфиопияда кең таралған келесі түршелермен едәуір дәрежеде).[28] Ересектер көбінесе осы номиналды кіші түрге қарағанда басқа нәсілдерге қарағанда салыстырмалы түрде қатты руфитке ие және кейде қараңғы жолақтармен ерекшеленеді. Сонымен қатар, кәмелетке толмаған жасөспірім болып келеді. аккорд осы жарыстағы ұзындық еркектерде 485-тен 540 мм-ге дейін (19,1-ден 21,3 дюймге дейін), ал әйелдердегі 509-ден 565 мм-ге дейін (20,0-ден 22,2 дюймге дейін) өлшенді.[3] Екі үлгідегі қанаттың аккордтың орташа ұзындығы A. r. рапакс ерлерде 501 және 512 мм (19,7 және 20,2 дюйм) және әйелдерде 541 және 545 мм (21,3 және 21,5 дюйм) өлшенді.[6][29] Екі жыныстың құйрығының ұзындығы A. r. рапакс өлшемдері 245-тен 295 мм-ге дейін (9,6-дан 11,6 дюймге дейін) а тарсус ұзындығы 79-дан 92 мм-ге дейін (3,1-ден 3,6 дюймге дейін). Дене массасы кем дегенде 36 бүркіттің жалпы сынамаларында шамамен 1,6-дан 3,1 кг-ға дейін болуы мүмкін (3,5-тен 6,8 фунтқа дейін).[3]
- A. r. белисариус; бұл жарыс тұрады батыс Африка дейін Эфиопия және оңтүстік-батысы Арабия солтүстіктегі оңтүстікке дейін Конго Демократиялық Республикасы және солтүстік Кения.[3][30] Бұл жарыс номинацияланған кіші түрлерге қарағанда «ұқыпты» болып көрінеді, мүмкін, бұл жарыстың жиі басқа нәсілдерге қарағанда шөл тәрізді жағдайларда жиі болып, осылайша қауырсындары тығыз болады.[3][31] Бұл номинациямен салыстырылғанда, көбінесе күңгірт және қоңырлау, жоғары реңк аз көрінеді. Сонымен қатар, жеке бозғылт морфы A. r. белисариус ақшыл номинацияға қарағанда көбінесе бір-екі көлеңке, бірақ олай емес. Біраз үлкенірек деп айтылғанымен, өлшеу деректері бұл кіші түрлерді үміткерлердің жарыстарына өте ұқсас мөлшерде көрсетеді. Еркектерде аккорд 495-тен 535 мм-ге дейін (19,5-тен 21,1 дюймге дейін), ал әйелдерде қанаттар аккорды 500-ден 562 мм-ге дейін (19,7-ден 22,1 дюймге дейін).[3] Хабарланғандай, қанаттардың аккордтарының орташа ұзындығы еркектерде 515 мм (20,3 дюйм) және 525 мм (20,7 дюйм) болды, бұл үміткерлер жарысына қарағанда сәл аз айқын жыныстық диморфизмді көрсетеді.[6] Дене массасына келетін болсақ, 1 еркектің салмағы 2 кг (4,4 фунт), ал үш аналықтың салмағы 1,9 - 2,5 кг (4,2 - 5,5 фунт) болды.[3][6]
- A. r. виндхиана; қоспағанда Арабия түбегі Бұл жарысқа Азиядан табылған барлық оңтүстік бүркіттер, мысалы оңтүстік-шығыста кіретін шығар Иран және Үнді субконтиненті.[5] Алайда, түсініксіздіктер қай жерде болатындығына байланысты A. r. белисариус аяқталады және виндхиана басталады, әсіресе Таяу Шығыс аудандар.[32][33] Кейде A. r. виндхиана толық түрлер ретінде ұсынылған.[34] Бұл кіші түрлер басқа нәсілдерге қарағанда орташа қараңғы және әдетте жылы руфустық тонға ие емес. Жалпы алғанда, ол реңкі жағынан дала бүркітіне ұқсас.[3] Ересек ирис кейде қоңыр болады A. r. виндхиана (тағы да дала бүркітін еске түсіреді). Бұл жарыстың бозғылт морфы африкалық тау бүркіттеріне қарағанда сұрғылт және онша рау емес, дегенмен, әдетте, кәмелетке толмағандар мен жетілмегендер кейде руфь болады. Бұл ең кіші кіші түрлер болуы мүмкін, бірақ тұтастай алғанда бүркіттің кең ауқымы бойынша өлшемдері өте аз өзгеретіндігі туралы куәландырады. Қанат аккорды өлшемдер ерлерде 473-тен 535 мм-ге дейін (18,6-дан 21,1 дюймге дейін) және 510-дан 560 мм-ге дейін (20-дан 22 дюймге дейін).[3][35][36] Еркектерде қанаттар аккорды шамамен 495 мм (19,5 дюйм), ал әйелдерде 525 мм (20,7 дюйм) құрайды.[6] Еркектерде құйрықтың ұзындығы 242-ден 258 мм-ге дейін (9,5-тен 10,2 дюймге дейін), ал әйелдерде 242-ден 285 мм-ге дейін (9,5-тен 11,2 дюймге дейін). The тарсус ер адамның ұзындығы A. r. виндхиана 80-ден 87 мм-ге дейін (3,1-ден 3,4 дюймге дейін), ал әйелдерде 84-тен 91 мм-ге дейін (3,3-тен 3,6 дюймге дейін).[35][36] Үндістандағы емделмеген ересектердің салмағы 1,5-тен 2,1 кг-ға дейін (3,3-тен 4,6 фунтқа дейін) болды.[5]
Сипаттама

Қарапайым бүркіт «талғампаз емес, қыруар көрінеді» деп саналады, бірақ оған тән аквилинді силуэт бар.[3] Бұл түрдің ұзын мойны және ұзын терең шоты бар, көзімен сызық сызығы, «саусақтары» бар орташа ұзын қанаттары және сәл дөңгелектенген шаршы және қысқартылған құйрығы бар, олар құйрықты еске түсіруі мүмкін басқа бүркіттерге қарағанда лашын. Аяқтардағы қауырсындар кең көлемді және үлкен көрінуі мүмкін.[3][11][37] Есепшот пен бас мықты және батыл, денесі пропорционалды, ал аяғы күшті, ал бет әлпеті қатты көрінеді.[11] Қоңыр бүркіт қонған кезде тіке отыруға ұмтылады, көбінесе дүмбілерде, бағаналарда, аласа ағаштарда немесе ағаштардың басында күннің ұзақ уақытында немесе көлденең қалыпта біршама тұрақсыз жүру үшін жерге түсуі мүмкін. Қанат ұштары отырғанда құйрықтың ұшымен бірдей болады.[3] Ересектерде сарыдан ақшыл қоңырға дейін сары-қоңырға дейін өзгеретін түрлі-түсті көздер болады, ал кәмелетке толмағандардың көздері қара-қоңыр болады. Жасы да, аяғы да барлық жаста сары болады.[3] Жалаңаш бүркіт полиморфты -де айтарлықтай жеке вариациямен түктер кейбіреулерді шатастыруы мүмкін, нәтижесінде кейде шаштың айырмашылықтары пайда болады.[3][5] Ересек жаста олар түрлі-түсті сұр-қоңырдан кейде жолақты (немесе одан да көп қарапайым) түлкі-руфустың және сары-сарыға дейін өзгеруі мүмкін.[3][38] Ересектердің көпшілігі әдетте жалпы сұр-қоңыр немесе қызыл-қоңыр түсті болып табылады, анда-санда бозғылт дақтар доғасында және іште жақын жерде көрінеді, жамылғылар денеге сәйкес біркелкі болады.[5] Ересектердің қауырсындары басқа қауырсындармен жиі бозарғанына қарамастан, желке үнемі қараңғы және біркелкі болады, ал басқаларында жиі кездесетін қарама-қарсы бозғылт қауырсындары жоқ Акила.[5] Әйелдер сәл үлкенірек болумен қатар, ерлерге қарағанда сәл қараңғы және жолақты болуы мүмкін.[3] Қара-қоңыр көбінесе Үндістанда кездеседі.[3][5] Ересектер көбінесе қанаттарының және құйрықтарының қауырсындарынан бөлек әр түрлі түстерді салыстырмалы түрде аз көрсетеді, алайда жаңа балқытылған үлкен қанаттар жамылғылары мен екіншілерде ұсақ ақшыл ұштар пайда болуы мүмкін, олар жабық қанат бойында ақшыл сызықтар құра алады, үстіңгі бөліктері ақшыл және ұшу қауырсындары бар.[3] Бастың денесі сияқты жиі ашық түсті болады, бірақ кейде қасы күңгірт, басқа жіңішке қоңыр жолақтар немесе қара иек көрінуі мүмкін.[3] Сонымен қатар, құйрық қарапайым немесе қараңғы торлы (шамамен 7 жұқа жолақпен). Қара морфты ересек адам негізінен қара, күңгірт қоңыр болады. Кейбір қара морфты бүркіттердің тозуы тұрақты емес жолақ немесе балқытатын қоңыр және қара қауырсындарды көрсетуі мүмкін.[3] Аралық морф қараңғы жалған Мантиямен және қанат жамылғымен жоғарыдан жоғары қоңыр түсті, әр түрлі жолақты немесе балқытылған ашық түсті, сондай-ақ басы тәжі немесе тәжі жақтары бозарған. Аралық морфтың астыңғы жағы едәуір руфитті (әсіресе Африканың оңтүстігінде) кеудесі мен қапталдары өте ауыр және кең жолақты қара қоңыр, бірақ кейде қарапайым қара шалбар мен криссумға қарама-қарсы қара қоңыр пайда болады.[3] Ересек бозғылт морфты бүркіттер әрдайым қараңғы ұшатын қауырсындар мен құйрықты көтеретін ақшыл дене мен қанаттар жамылғылары арасында айқын қарама-қайшылықты көрсетеді. Бозғылт морфтарда астыңғы жағы ақшыл-қоңыр-қоңырға дейін қыл-қыбырлы болып келеді, бірте-бірте қараңғыдан азырақ, ал ортаңғы қанат жамылғыларынан қара қоңырға дейін қара-қара түске дейін үлкен қара жамылғылар мен ұшатын қауырсындарға айналады. Бозғылт морфты бүркіттердің басы ақшыл болуы мүмкін, бірақ кейде жіңішке қоңыр жолақтары немесе қара иегі бар. Бозғылт морфты ересектердің астында ашық түсті немесе қоңыр түсті, кейде іш аймағынан төмен бозғылт түсті. Тозған бүркіттердің бозғылт морфының дене қауырсындары дерлік ақшыл болып көрінуі мүмкін.[3] Қараңғы морфты жасөспірімдердің қара бүркіттері, әдетте, ақшыл-қоңыр түсті және қоңыр-қоңырдан, төменгі артқы жағынан жоғарғы құйрық жамылғыларымен ерекшеленеді. Кәмелетке толмағандар ақшыл-қара түсті қоңыр түсті үлкен жамылғылар мен ремиктерді көрсетеді, ал құйрық сұр және қоңыр түсті, әдетте тар кремді ұшы бар. Қара морфты жасөспірімдер көбінесе күңгірт түстерге балқымас бұрын бозарған немесе кілегейге айналуы мүмкін. Кейінгі кезеңдер онша танымал емес, бірақ қара морфты субадальттар мантияға, сондай-ақ бас пен жоғарғы кеудеге артқы денені ұстай отырып (яғни бел және жамбас жамылғысы) қою қоңыр немесе руфусты қоңыр түсті біртіндеп көрсетеді. . Әдетте басқа морфтар ұқсас, бірақ онша танымал емес және жеке-жеке сәйкес келмейді. Көбісі балқымадан кейін қатты немесе құмды, бірақ кейінірек мылжыңға айналады, ақшыл қауырсындардың мөлшері олардың ересек морфының белгісі болуы мүмкін.[3]

Ұшып бара жатқанда, бүркіт ұзын мойынға айтарлықтай шығыңқы басы бар, терең кеудесі бар, ұзын және кең қанаттары бар, жеті саусақпен біршама тар қолымен үлкен рэптор болып көрінеді.[3] Қанаттың артқы шеті сыртқа қарай қисайып, праймерия мен секундар қосылысында шегініп тұрады, ал дөңгелектелген, ұзындығы орташа ұзындықтағы құйрықты жайып ұстайды.[11] Ілінген қанаттардың терең соққысы олардың ұшуын едәуір ауыр және баяу етіп көрсете алады, бірақ олар қанаттардың қозғалыстарында тезірек және кеңірек, көбінесе күштірек болып көрінеді. Акила дала қырандары сияқты және оларды тонау үшін басқа рэпторларды қуған кезде өте епті бола алады.[3][5] Тауан бүркіттері жалпақ қанаттарымен қалықтайды немесе өте сәл көтеріліп, қолдары сәл төменірек болады, олар сырғанақта да ұшуы мүмкін, бірақ жылдам сырғанақ кезінде тым доғалы болуы мүмкін.[3] Ересектерге арналған қара морфтар жоғарыда және төменде біркелкі қара-қоңыр түсті, екі жағында да айқын емес және сәл бозарған және сұрғылт праймерді көрсетеді. Жоғарыда қараңғы морфтарда негізгі контраст қараңғы кілегейлі жамау болып табылады, ал төменгі жағында сұр түске қара ұштармен және қанаттар мен құйрықтар бойымен диффузиялық артқы жиегімен қарама-қарсы қойылған. Аралық морфты бүркіттердің артқы жағында қоңырдан руфусқа дейін қоңыр жолақты және қанат жамылғыларында қараңғы морфтар сияқты жоғары қарама-қарсы бозғылт бөртпелермен әр түрлі руфисты болып келеді. Аралықтың қою қара жолақтары астында әр түрлі болады және олардың түсі біркелкі көрінуі мүмкін. Аралық морфтардың қанаттарының квадраттары көбінесе бозғылт, ішкі қарабайырлар мен қара қанаттардың ұштарының қарама-қайшылығы күшті болады. Бозғылт морф - бұл қанаттардың екі жағындағы ақшыл немесе бұталы, олар үлкен қара жамылғылар, ұшатын қауырсындар мен құйрықтар мен әдетте скапулярлар бойынша бөлінген қара қоңырға қатты қарама-қарсы келеді. Праймериздер бозғылт морфтарда едәуір бозғылт, кейде ақшыл карпальды үтір сияқты. Кейбір бозғылт ересектерде барлық төменгі суреттерге ақшыл негіздер болады, ал квиллдер кейде қоршалмайды, бірақ көбінесе қауырсындарда тығыз, бірақ тар қара жолақтар болады.[3][5] Қара морфты кәмелетке толмағандар - жеңіл реңді және бозғылт түсті денеге тән, олар қара-қоңыр түсті үлкен жамылғылармен, артқы қабыршақпен, ұшатын қауырсындармен және құйрықтармен қатты ерекшеленеді, бұл өз кезегінде қаймақ тәрізді белден құйрық жамылғыларға дейін көрсетеді. Қараңғы морфты ювенильдер ақшыл морфты ересектерге ұқсас болуы мүмкін, олар ақшыл жиектерден және көбінесе қанаттардың үлкен жамылғыларының ұштары бойымен тұрақты емес бозғылт диагональдардан тұрады, бірақ әдетте олар ерте сөнеді. Түстердің дамуы аз, бірақ жас бүркіттер моль қоңырға айналады, қанатты төсемдерде 1-3 қараңғы жолақты көрсететін аралықпен жамылғыш болады.[3] Ересектердің астыңғы жағы (яғни шамамен 2 жастан 3 жасқа дейінгі) әдетте екі түсті, көкірек, қарын және астыңғы жамылғылары қара қоңыр, ал астыңғы жағы кремді ақшыл түсті. Бұл екі тонды өрнек Үндістаннан да, Африкаға дейінгі субагенттік бүркіттерден де көрінеді.[5][39] Ересектердің түктері өмірдің 4-5-ші жылдары аралығында алынады.[3]
Өлшемі

Бұл үлкен жыртқыш құс, бірақ орташа өлшемді бүркіт және бұл тұқымдастың кішігірім түрлерінің бірі Акила. Қазіргі уақытта түрге қабылданған түрлердің арасында олардың мөлшері ұқсас Бонеллидің бүркіттері (Aquila fasciata) (дегенмен ұзын қанатты), қарағанда сәл үлкен Африкалық бүркіттер (Акила спилогастері) және қарағанда әлдеқайда үлкен Кассиннің бүркіттері (Aquila africanus). Әйтпесе, үлкен түрлерінің аналықтары Акила олар көбінесе орташа бүркіттен екі есе ауыр.[40] Жыртқыш құстарға тән болғандықтан, аналық бүркіт еркектерге қарағанда үлкенірек, дегенмен салыстырмалы түрде қарапайым және жыныстар арасындағы айырмашылық әдетте 15% -ке дейін жетеді.[3] Жалғыз бүркіттердің ұзындығы 58-ден 75 см-ге дейін болады (23-тен 30 дюймге дейін).[5][41] Жалаңаш бүркіттің ұзындығы шамамен 65 см (26 дюйм) деп саналады.[42] Қанаттар 157-ден 190 см-ге дейін (5 фут 2 футтан 6 фут 3 дюймге дейін) өлшей алады. Салмағы толығымен өсірілген құстарда 1,5-тен 3,1 кг-ға дейін болуы мүмкін (3,3-тен 6,8 фунт).[3][5][43] Орташа салмақтар бір зерттеуде 5 еркекте 1,91 кг (4,2 фунт) және 5 әйелде 1,97 кг (4,3 фунт) деп көрсетілген.[29] Тағы бір зерттеуде 10 ересек ересек бүркіттің салмағы орта есеппен 2,5 кг (5,5 фунт) екені анықталды, сол мәліметтер қоры үшін 15-дің орташа қанаттарының ұзындығы 182,9 см (6 фут 0 дюйм) болды.[44] Африка еркектерінің тағы бір кішігірім үлгісі, олардың төртеуі, орта есеппен 1,85 кг (4,1 фунт), ал үш аналықтары 2,28 кг (5,0 фунт) құрады.[6] Бір бағалау бойынша түрдің орташа массасы 2,3 кг (5,1 фунт) құрады.[45] Біріктірілген барлық стандартты өлшемдерде қанат хордасы 473-тен 565 мм-ге дейін (18,6-дан 22,2 дюймге дейін), құйрығы 242-ден 295 мм-ге дейін (9,5-тен 11,6 дюймге дейін) және тарсус 79-дан 92 мм-ге дейін (3,1-ден 3,6 дюймге дейін) өзгеруі мүмкін. .[3][46] The culmen Кениялық бүркіттердің ұзындығы 33,3-тен 42,4 мм-ге дейін өлшенді (1,31-ден 1,67 дюймге дейін), орта есеппен 37,7 мм (1,48 дюйм), ал ұшақтың ені орташа есеппен 44,3 (1,7 дюйм), 40,3-тен 49,9 мм-ге дейін (1,59-дан) 1,96 дюйм).[20][47] Акципитридтерде өлтіру құралы ретінде жиі қолданылатын галлю-тырнақ, өлшемі 27-ден 37,7 мм-ге дейін (1,06 - 1,48 дюйм) өлшенеді, бір үлгіде орташа алғанда 31,8 мм (1,25 дюйм) және 32,3 мм (1,27 дюйм) басқа. Бот бүркіт үшін талон мөлшері әсіресе үлкен емес және мөлшері бойынша дала бүркіттерімен пропорционалды түрде ұқсас шығыс империялық бүркіттер (Aquila heliaca).[6][20][47]
Шатасу түрлері

Жалаңаш бүркіт басқа жерлерде жиі кездесетін қоңыр қоңыр және үлкен рапторлар жиі кездесетін жерлерде өмір сүреді. Осылайша сәйкестендіру сирек қарапайым деп айтуға болады.[48][49] Оның қыстау аймағында ықтимал шатасушылықты тудыруы мүмкін бір көзі - бұл бұрыннан белгілі болды дала бүркіті. Дала мойны қысқа, қанаттары салыстырмалы түрде ұзын және тар, тұмсығы үлкен, әсіресе тұмсығының айрықша тереңдігі арқылы (егер ұшып бара жатқанда мойыны онша шығыңқы болмауы мүмкін) және ұзын әрі дөңгелек құйрығы бар. Сонымен қатар, дала бүркіттері бүркіттерге және қанаттардың айқын қараңғы шеттеріне және бозарған жұлдыруына қарағанда әлдеқайда батыл және кең аралықтағы тосқауылдарға ие.[3][20][49][38][50] Дала бүркіттерінен басқа симпатикалық бүркіттердің басқа топтарын салыстыру жүргізілуі мүмкін. Салыстырғанда дала бүркіттері, аң аулайтын бүркіттердің құйрығы ұзын, шоттары анағұрлым айқын, қанаттары әдетте төртбұрыш түрінде көрінбейді, мойны ұзын және түсі жалпыға ұқсас болып келеді. Айырмашылығы империялық қырандар, жалаңаш бүркіттің қанаттары кеңірек және артқы жиектері аз, есепшот сәл кем көрінеді және дене өлшемдері кішірек болған кезде қанаттар сәл жоғары қаралады. Бүркіт түрлерімен салыстырған кезде, олар тек алтын және алтын Верроның бүркіті (Aquila verreauxii) әдетте өзекті болады (осы эволюциялық тізбектің құрамында үш бірдей емес және симпатикалық аққұбалық бүркіттер, олардың мөлшері бүркіттері бар немесе одан кіші болса да, кездеседі), тау бүркіттің мөлшері едәуір кіші, қанаттары конустық емес олар күшті диедралда және пропорционалды түрде ұсталуы мүмкін емес пе, есеп айырысу ұзағырақ, ал құйрығы анағұрлым қысқа.[2][5] Үлкен ала бүркіттер (Кланга кланы), дала бүркіті сияқты а Палеарктика көбінесе жалаңаш бүркіттердің қыстауында қыстайтын өсіруші бүркіт өте ұқсас, бірақ бұл түрлердің құйрығы салыстырмалы түрде қысқа және кең, аяғында қылшық қауырсындар аз болады және ересектер түктерінде біршама қараңғы және біркелкі болады.[3] The фулвесцендер үлкен бүркіттің формасын ашық бүркіттің бозғылт түрлерінен олардың асты өрнектері арқылы, көбінесе қара түсті астарлық жамылғылармен және әдетте ашық түсті қара сазды доға арқылы бүкіл праймериздің үстінен қарапайым көрінетін қара реңктермен ажырату керек.[3][51] Көші-қон аз бүркіт (Кланга помарина) жалаңаш бүркіттен кішірек және құйрығының үстінде айқын U бар ақылды.[3][51] Африка тұрғын үйі Уолбергтің бүркіті (Hieraeetus wahlbergi) бүркіттегі сияқты біркелкі қылшыққа ие бола алады, бірақ әрқашан ұшатын қауырсындары сұрғылт және қанаттары салыстырмалы түрде ұзағырақ және тіктөртбұрышты және ұзын, тар және түзу құйрығы бар бүркіттерге қарағанда әлдеқайда аз.[3][51] The шығыс империялық бүркіт кәмелетке толмаған жасөспірімдерде бозғылт және аралық морфты қараңғы бүркіттерге ұқсас көрінуі мүмкін, бірақ императорлық бүркіт әдетте айқын, үлкенірек, жіңішке, ұзын қанаттарымен, ұзын, кең құйрығымен, сондай-ақ кеудесінде қара қоңыр жолақтармен, мантия мен қанат жамылғыларымен және ақшыл артқы жиектері мен қанаттарының сырықтары.[3][38] Үндістандағы қара морфты бүркіттерді осындай өлшемдерден ажыратуға болады қара бүркіттер (Ictinaetus malaiensis) соңғысы неғұрлым ұзын, күңгірт және қалақ тәрізді қанаттарымен жіңішке, негізі тар және құйрығы әлдеқайда ұзын, тар және айқын.[3] Сияқты бір-біріне ұқсамайтын бүркіттер Шіркеулер, яғни қоңыр жылан бүркіттері (Circaetus cinereus), жылан бүркіттері (Circaetus pectoralis) және кәмелетке толмаған бателистер (Terathopius ecaudatus), кейде шатасудың ықтимал көзі ретінде айтылады, бірақ, әдетте, бір-біріне ұқсамайтын реңктерінде де айтарлықтай ерекшеленеді (түгелдей үлкен, өте ұсақ шоқтығы бар, қысқа құйрықты және жалаң аяқты).[11][51]
Қоңыраулар
Тауан бүркіттері көбінесе үнсіз.[3] Алайда дала бүркіттерінен айырмашылығы, олар өздерінің өсіп-өнетін жерлерінен мүлдем үнсіз, кез-келген маусымда анда-санда дауыстайды.[38] Ол бүркіттерге қарағанда көбеймеген кезде дауысты болады.[3] Әдеттегі қоңырау - бұл әртүрлі, транскрипцияланған өте қатал, қуыс және қатты қабық коук-коук, кау-кау, киок-киок, ки-арк және тағы басқа. Шақыру өте жоғары (дала бүркітіне қарағанда сәл аз тереңірек), бірақ бүркіттерден гөрі тереңірек айтылады.[3] Жылы Крюгер ұлттық паркі бұл қоңырау қатты және алыс сапар деп айтылады.[11] Еркек бүркіттер - бұл түрлерде, әсіресе аспан билері кезінде, сонымен қатар басқа контексте жиі кездесетін вокалистер. Оларға тамақ таластары, ұя салу кезіндегі бұзушылықтар және еркектердің аналықтарын тамақтануға тартуы жатады.[3] Тоғыз жыл ішінде аң аулайтын бүркіттерді бақылау Зимбабве дегенмен, қоңырау бір рет естілмеді.[52] Оның үнсіздігінің себебі ол мекендеген тегіс ландшафтқа байланысты болуы мүмкін.[52] Жазылған басқа қоңырауға қатаң тор кіреді к еке ке... әуе кездесулерінде және жұтқыншақта кра клептопарастикалық ізденістерде. A кра-кра зиянкестерді ескерту үшін кейде қоңырау шығуы мүмкін.[3][5] Сондай-ақ, аналық безеуілдеуі мүмкін шрип-шрип ұяда, сондай-ақ сирек кездесетін айқай-шу (сәйкесінше тамақ сұрау және дабыл қағу мүмкін).[3][5][53] Жас балапан алғашқы бүркіт чиптері, бірақ қауырсындары пайда болғаннан кейін, ол қатты шақырумен жалбарынуға бейім, яғни. біз-йик, ви-йик.[11]
Таралу және тіршілік ету аймағы
Тауан бүркіттері өте кең таралған. Африка халқы үш бөлек популяцияда кездеседі.[3] Олардың біреуі табылған Солтүстік Африка оңтүстік-орталықта Марокко, мүмкін солтүстік Алжир, оңтүстік-батыс Мавритания, Сенегамбия, оңтүстік Мали, орталық және оңтүстік Нигер шығысқа қарай оңтүстікке қарай Чад, солтүстік және орталық Судан көпшілігіне Эфиопия және Сомали (бірақ солтүстік-шығысқа және орталық-шығысқа).[1][3][28][54][55][56][57] Солтүстік Африка халқы аз. Мароккода, олар кейбір аймақтарда қалған бірнеше популяциялармен қатты сарқылуда Тарфая, Таң-Тан және Соус-Масса.[58] Олар мүмкін жойылған бастап Тунис, олар бір кездері жиі болатын.[59] Жылы Батыс Африка, кейбір бүркіттер кездеседі Гамбия, Бару, Нигерия және (мүмкін, асыл тұқымды емес) Кот-д'Ивуар және Гана.[60][61][62][63][64][65][66] Жылы Африканың шығысы және орталық Африка, жалаң бүркіт орталық және шығыста кездеседі Конго Демократиялық Республикасы және құрғақ бөліктері бойынша Уганда және бүкіл халықтарда Кения, Танзания, Замбия (көбінесе Луангва алқабы және Чамбеши дренажы ), Малави және Мозамбик. Африканың шығысында бұл ең көп таралған және үнемі көретін қоңыр бүркіт болып саналады.[67][68][69][70] Жылы Африканың оңтүстігі, жалаңаш бүркіт барлық жерде кездеседі Зимбабве (қазір сирек кездеседі) Матабеландия және Чипинга таулары ), Ботсвана (әлі де тұрақты Окаванго атырауы ) және кейбір аймақтары Намибия, оңтүстік және батыс Ангола (Cuando Cubango, Кунене, Хила Намибе, дейін Малендже ), Эсватини, Лесото және солтүстік және орталық бөліктері Оңтүстік Африка, яғни негізінен солтүстігінде Апельсин өзені бірақ кейде Кейп провинциясы.[1][11][71][72][73][74][75][76][77][78] Таза бүркіт өсіруші ретінде жойылып кетуі мүмкін Свазиленд соңғы рет 2001 жылы өсірілгені расталды.[79]

Африкадан тыс түрлер, мүмкін, оңтүстік-батыс бөлігінде болуы мүмкін Арабия түбегі, яғни Йемен және шеткі оңтүстік-батысы Сауд Арабиясы ішінде Тихама және 'Asir аймақтары, бірақ соңғы бірнеше онжылдықта репродуктивті оқиғалар туралы расталған жоқ.[1][80][81] Жалаңаш бүркіт сирек қаңғыбас болып саналады Израиль кейбіреулері расталса да, олардың басқа хабарлары көбінесе дұрыс анықталмаған дала бүркіттері болып шығады.[82] Олар сондай-ақ сирек кездесетін қаңғыбас ретінде белгілі Оман.[83] Азияда аңшы бүркіт оңтүстік-шығыста оқшау тұрады Иран (Арабияда сияқты, тексерілген жақында өсіру белгісіз) және шығыста біршама үздіксіз Пәкістан (жиі Инд алқабы ), көп бөлігі солтүстік және түбек Үндістан, шығысқа әрең дегенде оңтүстік арқылы Непал және Ассам.[3][84][85][86] Непалдың бүркіттері сирек кездесетін болса да, бұл түрлер әлі күнге дейін ойпаттағы жартылай шөлдерде тұрады деп ойлайды.[87] Үнді диапазоны - бастап Пенджаб арқылы Үнді-Ганг жазығы және батыс Бенгалия, солтүстік-шығысы Бихар, Декан үстірті диапазонға дейін жалғасады Андхра-Прадеш, Карнатака және (негізінен солтүстік-орталық) Тамилнад.[5][84][88] Бүркіттердің айналасында қозғалатын жазбалар Мьянма, Солтүстік Вьетнам және Тайланд Дала бүркіттері дұрыс анықталмаған болуы мүмкін немесе қазір анықталмаған үлгілерге негізделген деп болжануда.[89] Кішкене бір қаңғыбастар келуге расталды Шри-Ланка (тек белгілі сыртқы түрі Акила бүркіт бар).[90] Растауды қажет ететін қаңғыбастық туралы ескі хабарламалар да белгілі Ауғанстан.[84]
Тіршілік ету ортасы

Тауан бүркіттері әр түрлі биіктікте ашық жерлерде кездеседі, бірақ әдетте тіршілік етеді құрғақ жерлер. Жылы Батыс Африка, түр көбінесе ылғалды көбейеді орман-саванна мозайкалары бірақ көшуге болады құрғақ орманды алқаптар және жартылай шөлдер асылдандырылмаған кезде.[3] Жылы Марокко, түр көреді орманды аймақтар жақын таулар іргелесімен жазықтар. Африканың басқа жерлерінде әдетте бүркіттер мекендейді орманды саванна сияқты құрғақ Акация саванна және жартылай шөл дейін шөлді аймақтар. Алайда, деревняные өсуі мүлдем жоқ экстремалды шөлді аймақтардан ылғалдан аулақ болуға болады тропикалық тропикалық ормандар. Сияқты кейде ол қолдан жасалған жерлерде кездеседі егістік жерлер, жол жиектері, бөгеттер, ауылшаруашылық жерлері, мал жайылымдары және ойын аймақтары егер оларда тамақтану мүмкіндіктері пайда болса.[3][28][70][74][75][91] Жылы Африканың оңтүстігі, тікенді көбінесе тірі бүркіттердің тіршілік ету ортасы болып саналатын тіршілік ету ортасы Акация. Ұқсас климаттық жағдайларға қарамастан миомбо орманы, жалаңаш бүркіт сирек кездеседі.[11][70][74] Үндістанда ұқсас тіршілік ету ортасы негізінен қолданылуы мүмкін, бірақ бүркіт көбінесе көбіне жақын жерлерде болуы мүмкін ауылдар және жиі өсіру қоқыс үйінділері және қасапханалар олар Африкадағыдан әлдеқайда көп.[3][5][84] Үндістанда құрғақ аймақтардың барлық градиенттерінен басқа, жалаңаш бүркіт жиі кездеседі тікенді ормандар.[5] Тауан бүркіттері теңіз деңгейінен 3000 метрге дейін тіршілік етуі мүмкін, бірақ біршама төмен биіктіктерді қалайды.[3] Бүркіттің мекендейтін мекендерінде күтілетін құрғақтықтың белгілі бір деңгейіне қарамастан, егер тіршілік ету ортасы белгілі бір сұранысты қанағаттандырмаса, олар әдетте ұя салмайды.[4] Таңғы бүркіттің болуы олардың болуына байланысты уақытша жауын-шашын кезінде ылғалды маусым.[4][92][93] Жауын-шашынның біршама тәуелділігі тіршілік ету ортасының сапасы мен жыртқыш популяциялардың белгілі бір дәрежеде болуы, сонымен қатар ұя салатын жерлердің болуы үшін маңызды болуы мүмкін.[11][92] Жалаңаш бүркіт, міндетті түрде, міндетті түрде ағаштың ұясы болып табылады және сол себепті ағаштың өсуіне қолдау көрсете алмайтын немесе ағаш өсетін жерлерде құрғақ болады. артық жиналды түрді сақтап қалуы екіталай.[5][11][94]
Мінез-құлық

Тау бүркіті, дала бүркітіне мүлдем ұқсамайды, негізінен отырықшы және миграциялық емес. Алайда Африкада оны кейде әділ деп санайды көшпелі және кейбір маусымдық қозғалыстармен айналыса алады.[3][95] Жылы батыс Африка, A. r. белисариус ылғалды орманды алқаптарға қысқа қашықтықты үнемі қазаннан қарашаға дейін жүріп, сәуірде солтүстікке оралады, мүмкін, кейде кем дегенде, Калахари аймағы туралы Ботсвана және оңтүстікке қыдыруы мүмкін Оңтүстік Африка.[3][62][64][96] Кейде жалаң бүркіттер Эфиопия мен батыс Африканың арасында жартылай жүйрік болып көрінеді.[95] Тіпті, қаңғыбас сияқты алыс қашықтыққа кезу туралы хабарланған A. r. белисариус жылы Тунис және қаншалықты Египет (мұнда 1950 жылдары екі рет жазылған) және тіпті Израиль (1990 ж. 3 қысқы жазба) және Оман.[3][97][98][99] Үндістаннан адамдар жақын маңда сирек кездеседі Бангладеш, бәлкім, дисперстен кейінгі кәмелетке толмағандардың қаңғыбастығы сияқты, бірақ түрлердің қаңғып жүргендігі туралы есептер Оңтүстік-Шығыс Азия сияқты Тайланд қазір апокрифтік деп саналады.[89][100] Сияқты салаларда, әдетте Африканың оңтүстігі, жабайы бүркіттер әдетте өздерінің өсіп-өнген жерлерінен сирек кездеседі және кәмелетке толмаған бүркіттер өздерінің ұяларынан бірнеше ондаған шақырымнан аспайды.[11][101] Ұя салатын құс Эсигодини жақын жерде қалпына келтірілді Форт-Риксон екі жылдан астам уақыттан кейін.[74] Алайда Африканың оңтүстігі үшін біршама алшақтықта ұя салған бір бүркіт төрт жылдан кейін Зимбабведе шыққан ұясынан 330 км (210 миль) қашықтықта қалпына келтірілді.[70] Табиғаттағы бүркіттердің бір-біріне сәйкес келмейтін және болжанбаған болып көрінетін қозғалыстары эксперименталды экологиялық зерттеулер арқылы дәлелденді, бұл бүркіттердің жаңбырдың жетіспеушілігін өтеу үшін жаңа аймақтарды іздеуінің мысалдары.[102] Дала қырандары көбінесе аздап өседі әлеуметтік және отар оппортунистік қоректену көздерінде жалаң бүркіт әдетте жалғыз болып саналады.[3][5] Алайда, кейде екі-үш бүркіттің топтары кездеседі, мысалы, Үнді субконтинентінде, бірақ кейде топтық өлшемдер бұл көрсеткіштен асып түсуі мүмкін.[5] Ішінде Мирпур дивизиясы туралы Азад Кашмир Пәкістанда үш жыл бойы оқыған бүркіттердің ұсақ топтары қараша мен ақпан аралығында жылы жерлерде жиналғанына куә болды.[103] Кішкентай топтар немесе жиынтықтар Африкада, сондай-ақ шоғырланған тағамдардың жанында пайда болатыны белгілі, тіпті қауымдық ағаштар ағаштарда, электр бағаналарында немесе жерде кездеседі.[3] Көптеген ірі рапторлар сияқты, жалаңаш бүркіт күнінің көп бөлігін қонуға жұмсайды, бірақ күніне бірнеше рет қанат алады.[5][11] Көптеген ірі бүркіттерден айырмашылығы, Үндістанда, кем дегенде, қанатты бүркіттер адамдарға әбден үйреніп алған және бақылаушылардың бір-біріне жақын орналасуына мүмкіндік беруі мүмкін.[5][104]
Азықтандыру

Жалаңаш бүркіт теңдесі жоқ Акила бүркіт оның тамақтану тәртібінде айқын мамандандырылмағандығында.[3][5][11] Басқа жағынан Акила оппортунистік тұрғыдан қиратады, жалаң бүркіт еркін жүреді қоқыс шығару қосулы өлексе, мүмкін, мұны жылдың кез келген уақытында жасайды.[5][11] Кәдімгі қоқыс тазалау көбінесе бүркіттерге жетелейді үйінділерден бас тарту ішінде немесе жанында ауылдар және қасапханалар, әсіресе Үндістанда және онымен жиі араласуға болады лашындар өлік алаңдарында. Олар сондай-ақ жол жиектерінде жиі жазылады жол жинағы тұрақты тамақ көзін ұсынады.[3][5] Кез-келген басқа рэпторға қарағанда әдеттегідей, бұл өте шебер қарақшы, үнемі басқалардың клептопаразитизмімен айналысады жыртқыш құстар.[5][11] Алайда, бүркітті «жалқау», «онша ерекшеленбейтін» және «әсер етпейтін» деп сипаттау онша орынды емес, өйткені бүркіт - өте үлкен жыртқыш аңдарды қоса алғанда, өте өзгермелі тірі жыртқыштарға шабуыл жасайтын өте қатал жыртқыш.[11][20] Бұл түр негізінен қысқа сүңгіп немесе алабұғадан секіру арқылы немесе қалықтаған ұшуда жоғарыдан еңкейіп аң аулайды.[3] Үнді субконтинентінде аңшылық алабұғаға артықшылық берілді Vachellia nilotica, Prosopis cineraria және Capparis decidua.[5] Сондай-ақ, ол көбінесе жерде жүру арқылы қоректенуі мүмкін. Жалаңаш бүркіттер көбінесе жерде жатқан тірі жыртқышты нысанаға алады, сирек нысанаға алады ағаш олжа.[3][5][11] Алайда, олар кейде ұшып түсіп, қанаттарға құстарды алады.[5][11] Әуеде өлтірілгені белгілі құстардың жыртқыштары ала көгершіндер (Колумба Гвинеясы) дейін фламинго.[11] Тауық бүркіттер көбейту кезеңінде жұппен жиі аң аулауы мүмкін, көбінесе көбейетін кезге қарағанда үлкен олжасын алады.[5] Кейде бұл тандем аң аулау жұп бойынша кез келген маусымда болуы мүмкін.[11] It is likely that tandem hunting by pairs involves one bird engaging in conspicuous flight to distract the quarry while another flies inconspicuously to sneak up on and kill the prey, as has been reported in other tandem hunting raptors.[5][11][105] Nocturnal animals сияқты гендер және springhares have been preyed upon by tawny eagles in areas where there was no possibility they were killed by traffic at night. In addition to other observations have been made where tawny eagles drink and bath at night, some nocturnal subsistence behaviour by this species has been inferred but no irrefutable evidence has been brought forth either.[11][106] Semi-regular attendance at grassfires in India, presumably in order to capture displaced creatures, has been reported.[84] More than 200 species, including both live prey and carrion, are known to be eaten by tawny eagles and they may have one of the most variable diet of all tropical eagles.[5][11][47][52][107][108] Reportedly, most prey the tawny eagle will take alive will weigh not less than 125 g (4.4 oz) and not more than 2.5 kg (5.5 lb), however live prey has been revealed to be regularly more variable than even that estimate represents.[3][11][47][52] One compilation study showed that, compared to 8 other Акила and spotted eagles, the tawny eagle's diet was the most evenly spread across all weight classes of prey from under 63 g (2.2 oz) to over 4 kg (8.8 lb), though took prey in the latter prey class slightly less so than the much larger алтын және құйрықты бүркіттер (Aquila audax). This study further determine that the most often focused on weight class in tawny eagle's diets were 0.5 to 1 kg (1.1 to 2.2 lb) and 1 to 2 kg (2.2 to 4.4 lb) prey class, accounting for a little less than half of the prey by quantity.[17]
Probable live prey

Determining whether prey has been taken alive at the nests of tawny eagles is generally considered to be difficult, although observations suggest that during breeding tawny eagles usually deliver fresh prey while raising young, indicating that such prey are usually either taken alive or newly pirated from other predators.[11][52][107] Within a Kenyan study, only 1.9% of prey brought to tawny eagle nests was thought likely to be from carrion.[47] While hunting prey, it often takes a variety of prey usually focusing somewhat on small to medium-sized сүтқоректілер, usually medium-sized to large құстар and occasionally medium to large-sized бауырымен жорғалаушылар.[11][47][52] A diet analysis in Зимбабве indicated that among 160 prey items, 36.9% were mammals, 51.9% were birds, 10% were reptiles and 1.2% were amphibians.[52] The leading prey species here were helmeted guineafowl (Numida meleagris), at about 21% of the prey total, and скрабты қоян (Лепус саксатилисі), at about 16% of the total.[52] A similar dietary study conducted in Лочинвар ұлттық паркі, Замбия found a higher proportion of birds and amphibians (61.4% & 5.5% respectively), with a surprisingly number of су құстары being taken, largely the қызыл түйме (Fulica cristata) және African pygmy goose (Nettapus auritus).[107] The variation in diet between the preceding two study sites is due to differences in habitat and prey availability.[107] A very detailed study was conducted in Цаво шығыс ұлттық паркі, Кения, of the diet of the local tawny eagles over different years. Of 543 total prey items, 41.2% were mammals, 35.4% were birds, and 23.4% were reptiles and amphibians. Of these the most important prey was Кирктің дик-дикі (Madoqua kirkii) at 21.7% of the total, yellow-necked spurfowl (Pternistis leucoscepus) at 6.8% of the total, мүйізді қоян (Лепус капенсисі) және crested francolin (Dendroperdix sephaena) both at 6.3% and red-crested korhaan (Lophotis ruficrista) at 5.9%; additionally, unidentified жыландар constituted 21.3% of the foods.[47] The dik-dik is a small бөкен and is much larger than a tawny eagle. The tawny eagle certainly took dik-diks weighing up to 4 kg (8.8 lb) and possibly even 5 kg (11 lb), with this eagle taking about 80 dik-diks in Tsavo East each year.[47] In different areas of Оңтүстік Африка, small dietary studies determined the diet at nests to be highly variable. Ішінде Highveld, about 52% of 60 prey items were mammals, 45% were birds and a small amount were балық. Жылы Тимбавати және Klaserie, 63% of the diet was birds, 34% of it was mammals and 3% were reptiles. At Highveld, 25% of the diet consisted of yellow mongoose (Cynictis penicillata), 15% Cape ground squirrels (Xerus inauris) and 13% helmeted guineafowl. At Timbavati and Klaserie, various франколиндер were strongly predominant in foods, at about 44% with another 17% by various монғулар.[109][110]
Out of the southern and eastern areas of Africa, less quantitative analysis has been undertaken into the feeding habits of tawny eagles, even around nests. What is known of their prey elsewhere is mainly from wide-ranging surveys, secondary accounts and photographs.[5][11] Ол пайда болады Эфиопия that the tawny eagle may have a close predatory relationship with the Абиссин шөптері (Arvicanthis abyssinicus) while one tawny eagle there tore open the nest of the Стресеманның бұтасы (Zavattariornis stresemanni) to access the prey.[28][111][30] Unidentified large rats constituted a great majority of prey delivered during the nestling growth stage at some east African nests.[112] Fewer details are known about the prey of tawny eagles in the Indian subcontinent.[5] One study, without quantitative data known, listed the prey of tawny eagles in Саураштра сияқты Үнді пальмасының тиіндері (Funambulus palmarum), eggs of red-wattled lapwings (Vanellus indicus), crows, монғулар, Indian hares (Лепус нигриколлисі) және тіпті Бенгал түлкісі (Vulpes bengalensis).[113] Otherwise, the prey incidentally reported in India is extremely varied, including even the hindlegs of a джунгли мысық (Фелис хаус) (but this may have been scavenged).[5] In general, tawny eagles in south Asia may focus on less varying prey in general, often reportedly favoring desert-dwelling кеміргіштер and hares.[84][114][115] Tawny eagles may hunt a couple dozen species of кеміргіш from different parts of the range, ranging in size from the 30 g (1.1 oz) Натальды мультимамматтық тышқан (Mastomys natalensis) to the 3.6 kg (7.9 lb) South African springhare (Pedestes capensis).[114][115][116] Жартас (Procavia capensis) және yellow-spotted rock hyrax (Heterohyrax brucei) are occasionally preyed upon by tawny eagles.[117][118] The tawny eagle may be one of the most accomplished predators of mongoose, many food studies reflecting relatively high numbers of them and they appear to be one of the most feared predators at мееркат (Suricata suricatta) colonies.[28][74][119][120][121] Ішінде Кароо, the local tawny eagles reportedly live mostly off of mongooses, either meerkats or yellow mongooses.[122] Similarly small or slightly larger жыртқыштар сияқты striped polecats (Ictonyx striatus) және гендер are not infrequently prey for tawny eagles as well.[11][109] Bat-eared foxes (Otocyon megalotis) may too be taken alive at times.[123] Әр түрлі маймылдар may be eaten, although not infrequently as carrion, tawny eagles may too in seldom cases attack juveniles of monkeys such as Patas monkeys (Erythrocebus patas), қайғы-қасірет (Хлороцебус этиопы) және маймылдар (Chlorocebus pygerythrus) up to the size of juveniles of several species of бабун.[47][117][124][125][126] However, unlike with larger eagles, the troops of certain baboons do not seem to regard tawny eagles as a threat based on their behavioural responses.[127] While most ungulate prey other than dik-diks is probably largely scavenged as carrion or stolen from other predators, the small calves of ungulates such as Томсонның газелі (Eudorcas thomsonii) are sometimes apparently killed by tawny eagles.[128] A general picture appears to emerge that tawny eagles quite often takes relatively large mammalian prey, surprisingly often creatures weighing up to 3 to 4 kg (6.6 to 8.8 lb) such as hares, dik-diks, the young of other antelopes, hyraxes and so on.[11][20][52][129]

While mammals prey varies from rodents to hares, mongooses and small antelopes, the diversity and size range of bird taken may be even more impressive and more than 120 avian prey species have been reported in the prey spectrum.[5][11][47][107][109] Included in the prey spectrum are various species of үйректер and small or gosling қаздар,[47][109][130] аңшы құстар, әсіресе франколиндер және теңіз құстары,[5][46][47][52] көп көгершіндер және көгершіндер,[11][35][131] mostly medium-sized species of құлақ және мүйіз[46][47][35][109][132][133] және көптеген су құстары from small курсорлар, лепвингтер, rails және гректер үлкенге фламинго, лайықты және бүркіттер both small and large.[11][47][109] Both young and adult Old World flamingoes of both African species are known to be attacked on occasion, as well as white-breasted cormorants (Phalacrocorax lucidus) және керемет корморанттар (Phalacrocorax carbo), all birds similar in size or somewhat heavier than the tawny eagle itself.[134][135][136] Even larger avian prey are taken including a кран (Grus grus) killed by a pair in Saurashtra (although it was an injured one) and presumably adult female Kori bustard (Ardeotis kori). If average-sized, these prey items likely weighed more than 5 kg (11 lb).[113][40][137] More minor avian prey includes түнгі машиналар, coucals, құм құмыра, жылдамдықтар, bee-eaters, балықшылар, біліктер, wood hoopoes, туракос, тотықұстар және бірнеше пассериндер.[5][11][35][47][109][138][139][140][141] One small passerine the tawny eagle may routinely hunt is the super-abundant қызыл есепшөп (Quelea quelea).[142] Жақын құс фабрикалары, tawny eagles can take to not infrequently lifting free-range тауықтар (Gallus gallus domesticus) және басқа да құс еті, especially when the eagles must feed their young, thus in turn potentially drawing ire of local farmers.[5][11][37][143]
Various snakes are taken opportunistically by tawny eagles and they can be quite bold about hunting улы жыландар.[144] In southern Africa and Kenya the following snakes have been identified in the foods of tawny eagles: Egyptian sand boa (Gongylophis colubrinus), young Африка рок-питоны (Python sebae), speckled sand racer (Psammophis punctulatus), rufous beaked snake (Rhamphiophis oxyrhynchus), қара мойынды түкіретін кобра (Naja nigricollis), қара мамба (Dendroaspis polylepis), бумсленг (Dispholidus typus) және қатпарлы қоспа (Bitis arietans).[11][47][145] They also hunt кесірткелер not infrequently given the chance, usually favoring fairly large species but capable of taking those ranging from геккондар дейін rock monitors (Varanus albigularis).[11][35] At one nest in Zimbabwe, кесірткелерді бақылау made up 29% of 83 prey items, but they were only 8% of 107 prey items of 3 other nests in the same park.[52] One of the most frequently seen prey to be taken by tawny eagles in India have been Indian spiny-tailed lizard (Saara hardwickii).[5][146][147] More minor prey have included тасбақалар, бақалар және құрбақалар және балық. A tawny eagle in southern Africa was seen to wade into shallow water and successfully pull out a largish лақа.[11][107][109][148] Tawny eagles can also take communal nesting and swarming insects fairly frequently. These are generally termites, which can attract several of these eagles especially amongst non-breeding eagles and young ones. When visiting termites, the tawny eagles commonly eat алатес and may, with an unusual lack of aggression, share the food source with several other birds of prey, including as many as a half dozen conspecifics.[149][150] At times, tawny eagles can also be attracted to swarms of шегірткелер.[151] In one case, a tawny eagle was seen consuming the fruit of an Адансония tree, an unusual instance of frugivory which is very rare in accipitrids other than one unusual species: the пальма жаңғағы (Gypohierax angolensis).[11] A tawny eagle was once witnessed picking through піл тезек бірге лашын, presumably searching for тезек қоңыздары тұтыну.[152]
Өлек

Although the tawny eagle does hunt for food, it also relies extensively on өлексе тамақ көзі ретінде.[11] Although most booted eagles and Акила will opportunistically feed on carrion, none is known to do so as routinely as the tawny eagle.[11][45] They have been recorded feeding on a huge array of carcasses as large as African bush elephants (Loxodonta africana) and at least as small as маймылдар and perhaps even down to the size of a көгершін.[11][153] Perhaps most frequently in Africa, tawny eagles will feed at carcasses of тұяқтылар сияқты бөкен.[11][52] At least 30-40 different species of ungulate have been recorded as carrion food-sources for these eagles.[5][11][47][52] At “vulture restaurants” in Эфиопия, feeding stations with dead livestock meant to mitigate the rapid decline in population of most African vultures, the tawny eagle was the second most often recorded scavenger at just under 35% of 1088 of recorded birds to feed at them.[154] The tawny eagle shares its carrion food sources almost invariably with лашындар and usually with several other scavengers such as шакалдар және гиеналар. Other birds that frequently also attend carrion are bateleurs, many other бүркіттер (оның ішінде steppe eagles ) және марабу ләйлектері (Leptoptilos crumenifer).[11][155][156] The producer-scrounger theory predicts that vultures rely on eagles for information on carcasses.[157] Due to their smaller size, eagles, i.e. tawny eagles and bateleurs, are able to begin foraging earlier in the morning and are thus more likely to locate a carcass first.[157] In 91 observed carcass in southern Africa, tawny eagles were verified to be the first to find 5 of them. Furthermore, vultures usually arrived in less than 40 minutes (in 75% of cases) after the tawny eagles found the carcass.[158] At large carcasses, there is a hierarchical social structure based on the size of the scavenger.[155] At Маасай Мара, the top scavengers were the considerably to slightly heavier mammals, i.e. spotted hyenas (Crocuta croctua), қара арқалы шакалдар (Canis mesomelas) және жабайы иттер (Canis lupus таныс), содан кейін лаппетке қараған лашын (Torgos tracheliotos), Рюпеллдің грифоны (Сыпандар rueppellii), followed by all other vultures with the tawny eagle and the bateleur in the second most and the most subordinate scavenger positions.[159] Similar scavenger hierarchies have been reported elsewhere as well.[155][156] Bateleurs were the most likely to first find a carcass of the Maasai Mara scavengers and both the tawny and bateleur were considered as scavengers with “low competitive ability and high search efficiency”.[159]
However, tawny eagles will at times be able to displace the smaller species of vulture such as капюшонды лашындар (Монахус некрозирттері) және Египет қарақұйрықтары (Neophron percnopterus), both of which are similar in weight to the tawny eagles themselves, with one tawny eagle even reportedly keeping as many as 20 vultures at bay at a carcass. In general, based on the literature, such an event of aggressiveness by this species at a large carcass would surely be unusual.[11][52][159][160] Tawny eagles do tend to be dominant over батель at carcasses, however.[11] Сығандар or griffon vultures are usually the most numerous vultures in attendance at carrion and are considerably larger than tawny eagles but sometimes may briefly tolerate a tawny eagle to feed in their midst depending on the circumstances.[11][161][153] Usually, the larger the group is of griffon vultures, the less likely the tawny eagle is to get to feed.[45][159][161][153] The eagles not infrequently remain on the periphery of the vulture feeding frenzy and wait for pieces of flesh to appear. Often they will be able to pick up small scraps but will wait until the carcass is finished and few vultures remain to feed. The tawny eagle can benefit from leading other scavengers to carrion or feeding subsequently to them since, unlike the largest and most aggressive vultures, such as лаппетке қараған аққулар және қарақұйрықтар (Aegypius monachus), the tawny eagle cannot tear open large carcasses on their own and tend rely on another source to access any bits of the nutritious viscera.[5][11][45][52][155][161] The tawny eagles when finding an unopened large carcass have few feeding options although may eat the eyes in such circumstances, as was verified in the circumstance of a tawny eagle finding a жылқы (Equus ferus caballus) carcass offered by researchers.[158] Roadkills are another feeding option as they are often torn asunder by impact with automobiles and the eagle may be able to (at least briefly) monopolize the carcass.[5][11][27] Perhaps not coincidentally, in Maasai Mara, the tawny eagles were found to benefit from a carcass being nearer human habitations and in lower quality habitats relative to the other scavengers.[159] In particularly in India, scavenging tawny eagles tend to regularly occur at полигондар where vultures seldom come but wintering steppe eagles may often feed alongside them seasonally.[5][35] Garbage dumps are also visited in different parts of Africa such as Уганда және Эфиопия by hungry tawny eagles.[162][163] Semi-predaceous and aggressively disposed vultures, like ақбас қарғалар (Trigonoceps oksipitalis) in Africa and қызылбас қасқырлар (Sacrogyps calvus) in India as well as the lappet-faced and cinereous vultures, tend to have little tolerance for tawny eagles, with the latter unlikely to approach until these aggressive vultures have had their fill.[5][11][52][164] On the contrary, though, at times white-headed vultures and tawny eagles have been observed peaceably sharing roadkills in some instances.[11] Often tawny eagles will come to smaller carcasses of almost any animal, as will other smaller scavengers like bateleurs and hooded vultures as well as crows, perhaps merely to avoid the competition that often occurs at large carcasses.[5][45][155][158][159] One subadult tawny eagle was observed to be following a pack of Африка жабайы иттері (Lycaon pictus), almost certainly in order to scavenge off of their kills.[165]
Клептопаразитизм

The tawny eagle steals food from other raptors in addition to catching its own prey and coming to previously dead food sources.[7][11] The Afrikaans name for the tawny eagle is a "Roofarend", meaning the "Қарақшы Eagle".[7] This behaviour is not entirely segregated from their scavenging on carrion behaviours but the considerable aggressiveness and boldness of the eagles in this circumstances are very different from their rather retiring disposition in scavenging contexts.[11][52][166] At times the tawny eagle is considered “fearless” in their piratical attacks and is certain to engage in them more frequently than almost any other member of the booted eagle clan or perhaps even birds of prey.[6][11] Other related eagles like the steppe eagle and eastern imperial eagle, as well as most теңіз бүркіттері, can be locally regular kleptoparasites but tawny eagles rob prey from other birds with some regularity in every part of the range.[5][11][46] Amongst all birds, only a few types of теңіз құсы сияқты скуалар және фрегат құстары are likely to derive a majority of their subsistence from kleptoparasitism.[167][168] Generally, tawny eagles will surprise other birds of prey with a dashing stoop and yank away the prey item in a manner of seconds; they will seldom completely land if the prey item is intercepted on the ground so they can take off with the plundered item quickly.[11][52][145] The size of birds that the tawny eagles have been known to pirate food away from have ranged from species as small as black-winged kites (Elanus caeruleus) және қарапайым қарақұйрықтар (Falco tinnunculus) to those as large as a lammergeier (Gypaetus barbatus).[6] There seems to few limits to the raptorial birds that the tawny eagle will not pirate from given the opportunity.[11][46] In one case, a pair of tawny eagles descended on a хатшы (Sagittarius serpentarius) that had killed a large қатпарлы қоспа and displaced both the secretarybird and an Африкалық қарақұйрық (Polyboroides типусы) that had tried to enter the fray, after which the eagle pair split the adder between them.[112] Other raptors known to be attacked for piracy in well-known and often repeated instances have included dark chanting goshawks (Melierax метаболиттері), bateleurs, lanner falcons (Falco biarmicus) and even the imposing әскери бүркіттер (Polemaetus bellicosus) және Верроның бүркіттері, the latter eagles having appeared to offer surprisingly little to no contest the tawny eagle's piracy despite their great strength and formidable talons.[11][52][169][170] Carnivorous birds that are not traditionally considered raptorial birds, such as марабу ләйлектері және southern ground hornbills (Bucorvus leadbeateri), are also occasionally kleptoparasitized by tawny eagles.[11][171][172] Interspecific piracy may be most frequent on bateleur despite that species being similarly sized and powered as the tawny eagle. While 5 displacements of tawny eagles by bateleur were reported in a study on their interactions, 26 instances of tawny eagles displacing bateleurs were described, clearly far more.[173] Several smaller birds of prey were observed to be repeatedly robbed of their catches at a қызыл есепшөп colony, including queleas crippled but not killed by lanner falcons, although some of the maimed queleas were contested by jackals as well.[11][52]
On occasion, a tawny eagle will find itself on the losing end of a kleptoparasitic interaction. Somewhat larger eagles have been seen to displace tawny eagles off of prey. Оларға жатады Африка балықтарының бүркіттері (Haliaeetus vociferus), eastern imperial eagles and their cousins, steppe eagles.[5][11][174][175] African fish eagles and Pallas's fish eagles (Haliaeetus leucoryphus) in India both seem to take precedence over tawny eagles at shared feeding sources such as carrion sites and water bird nesting colonies.[6][5] Ішінде Бэйл таулары of Ethiopia, бүркіт appear to engage in displacement of and may dominate the much smaller tawny eagles.[176] As aforementioned, a bateleur can succeed in seldom instances in pirating tawny eagles.[173] Vultures, especially лаппетке қараған аққулар, may assert themselves at recent tawny eagle kills and certainly can displace the eagles in some circumstances; it is likely but not confirmed that шакалдар may too opportunistically rob eagles as they have been recorded doing with other eagles. Large kills, which can not infrequently include prey of up to twice the eagle's own weight, are beyond the tawny eagle's ability for қанатты жүктеу. Such kills are probably frequently lost to other carnivores.[155][159][145] In Ethiopia, Ethiopian wolves (Canis simensis) were seen to rob tawny eagles repeatedly of freshly-caught rodents, succeeding in 5 of 21 attempts to do so.[177] Even much smaller birds such as house crows (Corvus splendens) have been seen to successfully rob a tawny eagle of its prey.[178]
Interspecific predatory relationship

The tawny eagle occurrence in Africa and the Indian subcontinent places it in arguably two of the most competitive environments for birds of prey in the world.[5][11] In turn, the tawny eagle seems to adapt via a lack of specialization on any particular prey type, hunting style or food source, and via including carrion in the diet quite often.[11][52] Many other raptors and eagles overlap in habitat use with tawny eagles.[7][11] However, wintering and residential Акила and spotted eagles that bear some relation usually use slightly differing habitats in contrast to the tawny eagle.[3][5][47] Жылы Цаво шығыс ұлттық паркі, the ecology of this eagle was studied at length in contrast to bateleurs and much larger martial eagles, which can appear to have broadly similar habitat and prey preferences, as well as the slightly smaller but larger-clawed African hawk eagles, which tends to habituate to slightly more wooded dry areas.[47] In general within this study, all four eagle species derived a majority of their prey biomass from Кирктің дик-дикі but that the martial and African hawk-eagles tended to take slightly larger dik-diks than the bateleur and tawny eagles, took slightly more in the park per pair based on annual estimates and were more unlikely to scavenge the prey.[47] The diet of the tawny eagle and bateleur in Tsavo East overlapped by around 64%, whereas the diet of the tawny and martial eagles only overlapped by 29%. The tawny eagle was the only eagle here to heavily supplement their diet with alternate prey like snakes, although bateleurs also took a wide range of prey.[47] The Tsavo East study further indicated that the predatory pressure on dik-diks is mitigated temporally by the slightly staggered nesting seasons of each eagle, with bateleurs tending to nest rather earlier, the hawk-eagle slightly later, so the peak reliance on the prey did not generally overlap.[47] Further study has indicated that, in Africa, the bateleur broadly mirrors the tawny eagle in most respects of ecology.[179] One stark difference from virtually any other known eagle is the tawny eagle's nesting habits. That is that this eagle nests almost invariably on the top of the canopy of a tree, rather than a main trunk or large sturdy branch of trees (or on cliffs or, in steppe eagles, the ground).[11][180] The nesting location of tawny eagles runs more parallel to those of лашындар.[155] Study in Крюгер ұлттық паркі has shown that the tawny eagle and white-backed vulture (Африкандық сығандар) will freely nest in the treetop nest built by the other species.[153] Furthermore, other species, including large үкі және жылан бүркіттері, will use old nests built by tawny eagles.[11] Although the habitats used by martial and tawny eagles have been reported as broadly similar, detailed study in the Кароо found that the tawny species preferred areas with higher and more predictable summer rainfall and with higher алғашқы өнімділік than the martial.[181]
Opportunistically, the tawny eagle will sometimes prey upon smaller birds of prey but this is fairly infrequent and the capture of raptorial birds has thus far been seldom reported. A hungry or food-gathering male tawny eagle may infrequently plunder the nests of other raptorial birds.[47][112] Incautious, injured or distracted birds of prey may too be vulnerable to being killed as well.[52][182] Diurnal birds of prey known to be preyed upon by tawny eagle in Africa have included black-winged kites, juvenile капюшонды лашындар, pale chanting goshawks (Melierax canorus) және African pygmy falcons (Polihierax semitorquatus).[47][46][109][182] In India, the tawny eagle has been known to prey upon western marsh harriers (Circus aeruginosus), shikras (Accipiter badius) және white-eyed buzzards (Butastur teesa).[35][113] Owls are apparently fairly vulnerable to tawny owl predation. Species that they have been known to prey on are қора үкі (Tyto alba), spotted eagle-owls (Bubo africanus), кішкентай үкі (Athene noctua), pearl-spotted owlets (Glaucidium perlatum) және marsh owls (Asio capensis).[47][182][183] The fresh remains of a хатшы were found in one tawny eagle nest in Africa but, if the eagles killed the bird rather than scavenged it, this would need confirmation.[11] The tawny eagle, despite being an eagle of intermediate size, does not seem to be subject to natural predators in adulthood as far as is known and can be said to fulfill the role of an шыңы жыртқыш.[11][46][107] Nests and young tawny eagle are commonly vulnerable to assorted natural predators but these are little known. A partial list of probable nest predators are likely коридорлар, жыландар және жыртқыштар capable of climbing.[3][52][107] One confirmed predator of nestling tawny eagles is the бал борсық (Mellivora capensis).[184]
Breeding biology
Pairing and Territories
The tawny eagle often seems to pair for life. Like most birds of prey, they are quite аумақтық towards conspecifics.[3][11] The commonest display is single or mutual high circling or soaring often in wide spiral. Males will sometimes dive and stoop repeatedly around the female, though she does not usually respond by turning over.[3] Pairs may engage in the display each to strengthen pair bonds.[5] Occasionally two tawny eagles will interlocks talons to descend rapidly, cartwheeling down 30 m or more within a few seconds, sometimes disengaging just before the ground.[5] In other related eagles of the Aquilinae subfamily, cartwheeling interactions are usually considered to be aggressive fights between a territorial eagle and an intruder of the same gender. Prior studies thought this to be the case for the tawny eagle, with an estimated 82% of cartwheeling instances thought to be aggressive, 11% for courtship and 7% for apparent play. However, through closer observations evidence has been made of frequent cartwheeling between males and females as a regular part of the courtship display.[185][186][187] Undulating sky dances are sometimes performed too by males with a series of descents and upward swoops on partially close wings, accompanied by calling. However, instances of this seem to be rare.[3] In one instance, two males appeared to engage in a display for a single female.[113] Per one author's opinion the aerial displays of the tawny eagle are “not particularly spectacular compared to other eagles”.[11] The breeding season tends to fall in March to August in northeastern Africa, October to June in west Africa and in almost all months of the year but in central, east and southern Africa, but mainly from May to November in Kenya and April to January in central and southern Africa.[3][188] In India, the breeding season is usually November to May, but occasionally can vary from any time from October to August.[3][5] Mating generally occurs in and around the nest vicinity.[5] The density very variable on the African continent overall of breeding pair which were estimated to occupy about 75 to 300 km2 (29 to 116 sq mi) each.[3] Zimbabwe nest spacing was found to be 7 to 10 km (4.3 to 6.2 mi) in one study. Шекарасында Крюгер ұлттық паркі, 7 pairs found in a 460 km2 (180 sq mi) area but in regular spaced pylon nests in western Transvaal, nests were 19 to 20 km (12 to 12 mi) apart.[11] Жылы Хванге ұлттық паркі, over 11 years of study, 92 pairs on were found to be nesting over базальт in a 4,724 km2 (1,824 sq mi) area while 84 pairs on Kalahari sands in a 9,876 km2 (3,813 sq mi) area. Mean nest distances on basalt were around 4 km (2.5 mi) while on sands it was around 59 km (37 mi).[4] Жылы Замбия, the nesting density was considered high for the species at a pair per 28 km2 (11 шаршы миль)[107]
Ұя
The nests of the tawny eagle are large platforms, composed of sticks but sometimes incorporating animal bones.[3] Nesting sites tend to be open to the sky, in flat, open or hilly country, and offer a commanding a good view of the surrounding country. The sites are not infrequently close to watering holes and, more so in India, close to villages.[3][5][11] Nests are usually 6 to 15 m (20 to 49 ft) above the ground, though seldom can be up to 30 m (98 ft) high.[3] Nests are located at the top crown of the tree and only very rarely are placed beneath the canopy or on a lateral branch.[3][11] Жылы Кения, tawny eagles showed no nesting preference according to tree height or spatial distribution of trees;[159] however, they preferred Эйфорбия, Боссия және Евклея ағаш түрлері.[159] In India, commonly used trees used in the northern areas are Ficus Religiosa, Dalbergia sissoo және манго trees while in the arid Kutch және батыс Раджастхан areas they often nest in rather stunted Vachellia nilotica және Prosopis chilensis (i.e. usually the nests here are 4 to 6 m (13 to 20 ft) high but are sometimes down to 4 m (13 ft) ).[5][189] Trees are usually selected that have prickly branches, presumably for protection.[11] Despite their prominent position in the trees, the nests can be surprisingly hard to perceive peering from the ground level.[11][188] Жылы Кгалагади трансшекаралық саябағы, Оңтүстік Африка, tawny eagles build nests that are positioned in the canopy of large Вахеллия эриолоба ағаштар.[93] These Kgalagadi pairs tend to be the largest and tallest trees, averaging at 10.9 m (36 ft).[93] Tawny eagles in India reportedly often nest in a tree over successive years, but the species is threatened by the lopping and cutting of all remaining suitable trees for fuel and fodder.[5] Tawny eagles in Kgalagadi Transfrontier Park, on the other hand, build new nests yearly and only 2% of nests are reused for breeding purposes the following year.[93] Potential risk of collapse and growth of branches around the nest are thought to be factors limiting nest reuses in Africa. Usually new nests are not more than 2 km (1.2 mi) away from the prior nest.[11] Екі жыныс ұя салуға қатысады және ұяны жөндеу 4-7 аптаға созылады, дегенмен құрылыстың көп бөлігі шамамен бір аптаның ішінде аяқталуы мүмкін.[11] Бүркіт үшін олардың ұялары салыстырмалы түрде кең, жалпақ және таяз.[11] Ұялардың диаметрі 1 м-ден (3,3 фут) және тереңдігі 20 см-ден төмен болуы мүмкін, бірақ қайта-қайта қолданған кезде 1,3 м (4,3 фут) пен 30 см-ден (12 дюйм) асады.[3][52] Ұялар әдетте шөптермен, жапырақтармен, тұқым бүршіктерімен және жүнмен, сондай-ақ газет, қағаз пакеттер және полиэтилен пакеттер сияқты тақ заттармен қапталған.[3][11] Жылы Крюгер ұлттық паркі, басқа бүркіттердің ұяларының көмегімен тіркелген рэптор сияқты аққұтан және ақбас лашын.[52] Кейде аң аулайтын бүркіттер ірі қауымдық ұялардың үстіне ұя салатыны белгілі болды ақбас буйвол тоқушы (Dinemellia dinemelli).[11] Орталықта Кароо аймақ Оңтүстік Африка, ұялы бүркіттер ұяларын үлкен электрлікке салады электр жеткізу мұнаралары.[190] Сияқты ірі бүркіттердің популяциясы әскери бүркіт және Верроның бүркіті 1970 жылдардан бастап осы қуат тіректерінде өсіру тіркелген.[190] 2002-2003 жылдар аралығында электр жеткізу желілерінде тіркелген электрлік ақаулардың 39% -ы үлкен бүркіт ұяларына байланысты болды.[190] Нәтижесінде, проблемалық ұялар бұзылып, төменде қайта салынды электр өткізгіштер.[190]
Жұмыртқа


Жұмыртқалар бірнеше күн аралығында, көбінесе құрғақ мезгілге, бірақ кейде ылғалды мезгілде басталады.[3] Дәлелдері Калахари шөлі жұмыртқалау жұмыртқалауды ұядағы балғындармен синхронды ауа райының жылы болуымен бірқатар тамақ ресурстарын пайдалануға, мысалы, әр түрлі ұсақ сүтқоректілер мен серпін (Antidorcas marsupialis) төлдеу маусымы.[191] Үндістанда өмір сүру ортасы онша оңтайлы болмаған кезде интервалдар ұзаққа созылды.[5] 1988 - 1996 жж. Аралығында бақыланған 26 бүркіт ұясының Кгалагади трансшекаралық саябағы, Төсеу күндерінің 84,6% мамыр мен маусым аралығында болды.[93] Бұл төсеу күндері популяцияларға ұқсас Замбия, Зимбабве [107][11] және Маасай Мара аймақ Кения сондай-ақ Африканың оңтүстік және оңтүстік-шығысындағы басқа жерлерде.[56][192][193] Африканың оңтүстігінде шілде және тамыз айларында жұмыртқа салатын жағдайлар іліністі ауыстыру жағдайлары болуы мүмкін.[11] Солтүстіктен және Батыс Африкадан фенология салудың кездейсоқ деректері бұл аймақтың аңшы бүркіттерінің жұмыртқаларын әдетте жылдың басында, яғни қаңтардан сәуірге дейін көрсетеді; Ганада жұмыртқалау қазаннан ақпанға дейін созылуы мүмкін, бірақ көбінесе желтоқсан-қаңтар айларында; Қараша-ақпан жұмыртқалары Эфиопияда, ал шамамен сәуірде Мароккода болды.[28][65][58][194][195][196] Үндістандағы вариацияны көрсету, in Пенджаб, Уттар-Прадеш және Бихар, бүркіттер негізінен қаңтарда жұмыртқа салады Кутч және Джайсалмер, жастар қазірдің өзінде ұядан кетіп жатыр.[5] Ілінісу мөлшері бір ұяға 1-ден 3 жұмыртқаға дейін, бірақ ілінісу үшін орташа есеппен 1,7 жұмыртқа құрайды.[11][107] Құрғақ жылдары Хванге ұлттық паркі, ілінісу өлшемдері кішірейтілген сияқты.[4] Жұмыртқалар ақ түсті, бірақ әр түрлі және әдетте әлсіз, қоңырмен белгіленеді, кейде белгісізден қызыл-қоңыр түсті дақтар мен дақтармен жақсы белгіленеді.[6] Ұсынылған кіші түрлердің 67 жұмыртқасында жұмыртқалардың биіктігі 64-тен 75,7 мм-ге дейін (2,52-ден 2,98 дюймге дейін), диаметрі 49,9 - 60 мм (1,96 - 2,36 дюйм), орташа 69,6 мм × 54,8 мм (2,74 дюйм) болды. Үлгіде 2,16 дюйм болса, сол жарыстағы тағы 30 адам 71,5 мм × 56,3 мм (2,81 дюйм 2,22 дюйм) құрады. Жылы A. r. виндхиана, 80 жұмыртқа 58-ден 75,1 мм-ге дейін (2,28-ден 2,96 дюймге дейін) 46,4-тен 57,6 мм-ге (1,83-тен 2,27 дюймге) дейін, орташа 66 мм × 52,8 мм-ге дейін (2,60 дюймде 2,06 дюйм).[6][11]
Жас және ата-аналық мінез-құлықты дамыту
Жұмыртқа инкубацияланған аналықтар 40-44 күн ішінде, 30-дан 45 күнге дейін шекті жазбалармен, балапан шығар алдында.[3] Инкубация бірінші жұмыртқадан басталады және оны тек Үндістандағы әйел жасай алады, бірақ африкалық мәліметтерге сәйкес, еркек кейде осында жеңілдейді.[3][5] Ұяларға көтерілгенде, аналықтар тығыз орналасқан, көбінесе соңғы минуттарда ұшып кетеді.[5] Жасыл төсем әлі де инкубация сатысында қосылуы мүмкін.[11] Жұмыртқадан шыққан кезде, жастарды үнемі ашық ұяларда өсіруге немесе күн сәулесінен көлеңкелеуге тура келеді.[3][6] Балапандар бастапқыда ақ мамықпен, қара шотпен, сары түсті аяқтармен және қоңыр көздермен жабылған; қалың ақ түсті пальто 2 аптада сатып алынады және 1 аптадан кейін қабыршақ пен қанат жамылғысында 1-ші қауырсын пайда болады.[11] Жас бүркіттер шамамен 3 аптада әлсіз тұра алады, 4 аптада ұяны айналып өтіп, шамамен бір аптадан кейін қанатты қақпалай бастайды.[11][52] Қанаттар мен құйрық жамылғылары тез көктеп шығады, қауырсындар 4 аптада емшек жағында пайда болады. 5-ші аптада қауырсындар дененің көп бөлігін бастан және астыңғы жақтан басқа жерлерді жауып тастайды. 7-ші аптада балапанның аз ғана мөлшері бар мамықтар қалған және салмағы 2,15 кг (4,7 фунт).[11][52] Дорсальды қауырсындардың қарқынды дамуы, мысалы, ашық ұяларды қолданатын басқа рапторлармен салыстыруға болады жылан бүркіттері және хатшы құстар.[11] Әдетте бір ғана балапан балапан шығарғаннан кейін тірі қалады.[11][46] Бұл көбінесе байланысты сиблицид, онда үлкен балапан өмірдің алғашқы бірнеше күнінде жас балапанды өліммен жаралайды. Үлкен бауырдың салмағы шамамен 143 г (5,0 унция), ал кішісі өлген немесе жоғалып кеткен кезде шамамен 85 г (3,0 унция).[11] Африканың оңтүстігінде бір ұяда екі жаңадан туылған кем дегенде төрт жағдай бар.[11][197] Шамамен 5 аптада бүркіт жыртқышқа қарсы әрекеттерді қабылдайды, кейбіреулері романға жақындағанда бейім, ал басқалары қауырсындарды көтеріп, тұмсықты ашып, қанаттарын ұруға дайын және талондарды кесуге тыйым салады.[11] Алғашқы 10 күнде ересек әйел еркек беретін тағамға сүйеніп, балапанды мұқият бақылайды. Екі аптадан кейін балапан күн сайын 2,5 сағат бойы жалғыз қалады, ал ересектер тамақтанады. Осы кезде ер адам бүркітке тікелей тамақ жеткізе бастайды. Бұл үлкен мөлшердегі бүркіттің ұясына баруды тоқтату үшін салыстырмалы түрде ерте кезең болып саналады, ал ұя көп ұзамай қалдықтармен жағымсыз болып қалады.[11][52] 39 күндік бүркітші өз тамағын жыртып үлгерді, бірақ оны бірінші кезекте ұрғашы тамақтандырды.[11] Алғашқы ұшу әрекеттері шамамен 7-10 апта, бірақ балапан толықтай өседі және қабілетті қашу 10-12 аптадан кейін ұя.[3][52] Алайда, ұрғашы жаңбырлы жаңбыр кезінде жаңа баспалдақ кезеңінде кешке дейін паналай алады.[3] Тәуелділіктің толық кезеңі қашып кеткеннен кейін шамамен 6 апта бойы жалғасуда. Жас бүркіт ата-анасында келесі өсіру маусымына дейін қалуы мүмкін.[3] Үндістанда, кем дегенде, ұя салу кезеңінен кейін жұптар тарайды және ұя салатын жерден кетеді, қазан айында жұптасу басталғанға дейін ұяның жанында сирек көрінеді.[5] 2 жасында атылған кәмелетке толмаған бүркіт өзінің ұясынан 48 км (30 миль) қашықтықта, ал 5 айлық және 7 айлық 2 кәмелетке толмағандар тиісінше 50 және 34 км (31 және 21 миль) қашықтықта болды.[11]
Сәттіліктер мен сәтсіздіктер
Жұмыртқалар мен жастардың ұядағы шығыны өте жоғары болып көрінеді.[3] Жас бүркіттер көбінесе, кейде олардың бауырлары өледі, ал егер нашар қорғалатын ұялардан жыртқыштардың кең ауқымы шықса.[3] Ұя салудың сәттілігі тіршілік ету ортасының сапасы мен тағамға қол жетімділікке байланысты.[5] Зимбабведегі асыл тұқымды күштер 26 жұпта 19 жас төл шығарды, олардың орнын ауыстыру коэффициенті жылына бір жұпқа 0,73 жас болды.[52] Үндістанда бүркіттердің жұптары субптимальды шамадан тыс құмды мекендерге бейімделіп, ұяларын көбірек шашыратады және соның салдарынан әр ұяға балапандардың өнімділігі ұқсас бола алады.[5] Хванге ұлттық саябағында бір жыл ішінде орташа есеппен жұптардың 72,4% -ы көбейеді деп ойланған, ал күш-жігерге орташа есеппен 0,61 балапан шығарылған.[198] Бұл және басқа зерттеулер жаңбырдың осы аймақтағы бүркіттерде жемісті жетістікке жетуінің кілті болып табылатындығын дәлелдейді, бір жұмыртқадан гөрі екі жұмыртқа ілінісі көп (көбінесе ол істен шыққан) және жауын-шашын көп болған жылдардағы өсу кезеңдері аз.[52][198] Жылына бір жұпқа жас ретінде жазылған асылдандыру жетістігі (ypy) әлі де төмен болды Намибия және Цаво шығыс ұлттық паркі қарағанда Зимбабве (Сәйкесінше 0,4, 0,5 және 0,78 ypy).[52][107] Ұя салудың жоғары жетістігі Замбияда табылды, мұнда жұп бір ұяға 1 жаңадан пайда болды.[107] Өмір сүру ұзақтығын зерттеуші жалаңаш бүркіт үшін жасалғаны белгілі болмаса да, бұл бүркіттердің табиғатта кем дегенде 16 жасқа дейін өмір сүре алатындығы белгілі.[7][199]

Күй
Сақтау
Тауық бүркіт әлі күнге дейін кең ауқымды алып жатыр.[1] Африкада бұл түрдің таралу аймағы шамамен 15 миллион шаршы шақырымды, Азиядағы шамамен 3,1 миллион шаршы шақырымды қамтиды деп есептелген.[3] 90-шы жылдары, жаһандық халықтың саны алты мың адам болуы мүмкін деп ойлаған, сол кездегі Азиядағы халқының саны жүз мыңға жетеді деп ойлаған.[3] Алайда, түр қазіргі уақытта тізімге енгізілген Осал үстінде IUCN Қауіп төнген түрлердің тізімі.[1] Қазіргі кездегі халық бұрын ойлаған санының жартысынан әлдеқайда аз, ал бүкіл әлемде шамамен 100,000-ден 500,000-ге дейінгі адамдар сақталады деп ойлайды.[1] Арасында бүркітті байқаудың төмендеуі байқалды SABAP және SABAP2 Африканың оңтүстігінде, торлы клеткалардың 1440-тың 323-інде ғана бар.[200] Орталықтағы әскери және әскери бүркітті мұқият зерттеу кезінде Намибия, екеуінде де айқын құлдырау анықталды, бүркіт популяциясы кезінде аймақтық саны 19 жұпқа дейін 2 белгілі жұпқа дейін болған.[201] Бұл түрдің бір кездері сансыз болып көрінетін популяциясы Кгалагади трансшекаралық саябағы 1990 жылдары белгілі 40 жұпқа дейін белгілі болды.[202] Жол бойындағы санау Мали, Нигер және Буркина-Фасо раптор түрлерінің көпшілігі популяцияның күрт азаюына ұшырағанымен, қорғалатын табиғи аумақтардан тыс жерде тек қана бүркіт пен жылан бүркіттері тіршілік етіп жатқанын көрсетіңіз.[203] Үндістанда жалаң бүркіт «біздің ең қарапайым бүркіт» болып саналды[37] деген сияқты бекіністерді көрсететін сауалнамалармен күшті құлдырау анықталды Раджастхан бақыланатын жұптардың жартысына дейін төмендеуін көрсетті.[5] Продюсер-скрунгердің жемшөптік теориясына сәйкес, лашындар белгілі бір дәрежеде ұшаларды табуға көмектесетін жалаңаш бүркіттерге сүйенеді.[157] Осылайша, бүркіттерді қорғалатын табиғи аумақтардан тыс жерде сақтау қарақұйрықтардың тіршілік етуін қамтамасыз ету үшін өте маңызды.[157]
Қауіп-қатер

Тауан бүркіттері олардың өсіру мінез-құлқына әсер ететін бірқатар қауіп-қатерлерге тап болады, жемшөпті өсіреді және сайып келгенде жеке құстардың тірі қалуына әсер етеді. Тіршілік үшін ең соңғы және жойқын қауіп 2019 жылдың 20 маусымында болды ұшалар 468 ақ сүйекті аққулар, 17 ақбас қарғалар, 28 капюшонды лашындар, 14 лаппетке қараған аққулар және 10 мүйізділер 2 бүркіттің жанынан табылды. Барлығы 537 лашындар солтүстігінде уланған 2 бүркіт табылды Ботсвана. Олар 3-тің өлігін жегеннен кейін қайтыс болды деген күдік бар пілдер браконьерлер улармен байлаған. Өлекселер браконьерлік жабайы табиғатты табу үшін табиғат қорғаушыларға көмектесе алмайтындығына көз жеткізу үшін уланады. Үлкен рапторлар өлген жануарлардың үстінен айналып өтіп, ерте анықтау жүйесі ретінде әрекет етеді браконьерлікке қарсы қорықшылар.[204][205][206][207] Улану оқиғалары тек Ботсванамен шектелмейді және олар бүркіттердің азаюының тікелей факторы деп саналады, тіпті қорғалатын аймақтарында Крюгер ұлттық паркі.[208] Орталықта Намибия, радиобелгіленген барлық 5 жасөспірім қанатты бүркіт стрихнин жемдерімен уланып, аймақтағы барлық түрлердің алынуын толығымен тоқтатты.[201] Жұмбақ күйде популярлар мен жалаңаш бүркіттер популяциясы Маасай Мара басқа жерлерде айтылған құлдырауға қарағанда, Маасайдағы құсбегілердің нашарлауымен үйлесетін сияқты.[209]
Қарапайым бүркіттер үшін бұдан кейінгі қауіп-қатерлерге тіршілік ету ортасын жоғалту және жерді пайдаланудың өзгеруі жатады, мысалы, малды жаюды күшейту, отын жинау және сату және көмір өнеркәсібі.[7] Құрғақ Үндістанның қосалқы ағаштарын осылай кесіп тастау Үндістандағы бүркіттердің аз түсінілуінің негізгі қозғаушысы болып көрінеді.[5] -Ның жоғары көрінісі бактериалды жұқпалы аурулар Үндістанның бүркіттеріне де әсер ететін сияқты.[210] Раптор популяциясы жыртқыш популяциялардың тіршілігіне әсер ететін маусымдық жауын-шашын оқиғаларына тәуелді.[7] Климаттық өзгеріс Оңтүстік Африканың құрғақ аймақтарындағы жауын-шашынның кезектесуі және жыртқыш популяцияларға әсер етуі. Жауын-шашын оқиғалары мен бүркіттердің өсу жетістіктері арасында нақты байланыс бар. Тау бүркіттерінің климаттың өзгеруінен болжанған құлдырауы, ол қазірдің өзінде популяцияның тұрақтылығы, алдымен популяция динамикасы, биологиялық қауымдастықтың құрамы және биоәртүрлілік әсер етуі арқылы басталатындығы анықталды.[7][211] Электр тоғымен және соқтығысу қаупімен байланысты әуе желілері үлкен бүркіттер үшін тұрақты қауіп болып қала береді және лашындар. Сонымен қатар, электр желісіне ұя салатын рапторлар маңызды көзі болып табылды желінің ақаулығы ауданда айтарлықтай қаржылық мәселелер туғызады.[190][212] Кейде аң аулайтын бүркіттерді де қолдан жасалған әртүрлі заттарға ұшып өлтіреді су қоймалары немесе болып табылады автомобильдермен қаза тапты және тәуекелге ұшырайды жел турбиналары Үндістанда[201][213][214] Кез-келген раптор популяциясының негізгі қаупі - бұл тұрғындардың көбеюі, бұл тіршілік ету ортасы мен тамақ ресурстарына бәсекелестік тудырады.[62] Бүркіт популяциясын сақтаудың кілті оның әсерін азайту болып табылады ғаламдық жылуы.[7][215][216][217][218] Сондай-ақ, бүркіт пайдаланылатын жерлерде улы жемге тыйым салу және қауіпті электр желілерін азайту - бұл бүркіттердің тірі қалуы үшін маңызды болып табылады.[201][205][190]
Әдебиеттер тізімі
- ^ а б в г. e f ж сағ BirdLife International (2018). "Акила рапаксы". IUCN Қауіп төнген түрлердің Қызыл Кітабы. 2018. Алынған 17 желтоқсан 2018.CS1 maint: ref = harv (сілтеме)
- ^ а б в г. e Helbig, A. J., Kocum, A., Seibold, I., & Braun, M. J. (2005). Акилиндік бүркіттердің көп генді филогенезі (Aves: Accipitriformes) экстенсивті парафилді тұқым деңгейінде анықтайды. Молекулярлық филогенетика және эволюция, 35 (1), 147-164.
- ^ а б в г. e f ж сағ мен j к л м n o б q р с т сен v w х ж з аа аб ак жарнама ае аф аг ах ai аж ақ әл мен ан ао ап ақ ар сияқты кезінде ау ав aw балта ай аз ба bb б.з.д. bd болуы бф bg бх би bj bk бл bm бн бо bp кв br bs bt бұл bv bw bx арқылы bz шамамен cb cc CD ce cf cg ш ci cj ck кл Фергюсон-Лис, Дж .; Кристи, Д. (2001). Әлемнің рапторлары. Хоутон Мифлин Харкурт. ISBN 0-618-12762-3.
- ^ а б в г. e Хастлер, К., және Хауэллс, В.В. (1989). Тропиктегі тіршілік ету ортасының артықшылығы, тұқымның жетістігі және Tawny Eagles Aquila рапаксындағы алғашқы өнімділіктің әсері. Ибис, 131 (1), 33-40.
- ^ а б в г. e f ж сағ мен j к л м n o б q р с т сен v w х ж з аа аб ак жарнама ае аф аг ах ai аж ақ әл мен ан ао ап ақ ар сияқты кезінде ау ав aw балта ай аз ба bb б.з.д. bd болуы бф bg бх би bj bk бл bm бн бо bp кв br Naoroji, R., & Schmitt, N. J. (2007). Үнді субконтинентінің жыртқыш құстары. Om Books International.
- ^ а б в г. e f ж сағ мен j к л м n o Браун, Лесли және Амадон, Дин (1986) Әлемдегі бүркіттер, сұңқарлар мен сұңқарлар. Wellfleet Press. ISBN 978-1555214722.
- ^ а б в г. e f ж сағ мен j Вихманн М .; Дин, В .; Jeltsch, F. (2004). «Жаһандық өзгерістер Tawny Eagle-ге қиындық туғызады (Aquila rapax): климат пен жерді пайдаланудағы болжамды өзгерістерге қатысты жойылу қаупін модельдеу". Түйеқұс. 75 (4): 204–210. дои:10.2989/00306520409485446.
- ^ Огада, Д.Л (2014). Удың күші: Африканың жабайы әлемін пестицидтермен уландыру. Нью-Йорк Ғылым академиясының жылнамалары, 1322 (1), 1-20.
- ^ Темминк, Дж. Дж. (1828). Үндістандағы Nederlandsche bezittingen компаниясының Sunda-eilanden-ге қызығушылығы бар. Биждр. нат. Ветенш, 3, 64-78.
- ^ Гилл, Фрэнк; Донскер, Дэвид, редакция. (2019). «Жаңа әлем қарақұйрықтары, хатшы, батпырауықтар, бүркіттер». Әлемдік құстар тізімінің 9.2 нұсқасы. Халықаралық орнитологтар одағы. Алынған 31 шілде 2019.
- ^ а б в г. e f ж сағ мен j к л м n o б q р с т сен v w х ж з аа аб ак жарнама ае аф аг ах ai аж ақ әл мен ан ао ап ақ ар сияқты кезінде ау ав aw балта ай аз ба bb б.з.д. bd болуы бф bg бх би bj bk бл bm бн бо bp кв br bs bt бұл bv bw bx арқылы bz шамамен cb cc CD ce cf cg ш ci cj ck кл см cn co cp cq кр cs Steyn, P. (1983). Африканың оңтүстігіндегі жыртқыш құстар: Оларды идентификациялау және өмір тарихы. Croom Helm, Бекенхэм (Ұлыбритания). 1983 ж.
- ^ Браун, Л. (1977). Әлем қырандары. Әлем кітаптары.
- ^ Li, Q., Lin, J., Li, S., Wang, Y., Li, W., & Zeng, Y. (2000). Accipitridae-дің 11 түріндегі митохондриялық ДНҚ эволюциясы туралы зерттеулер. Dong wu xue bao. [Acta zoologica Sinica], 46 (2), 209-220.
- ^ De Boer, L. E. M., & Sinoo, R. P. (1984). Accipitridae (Aves: Falconiformes) кариологиялық зерттеуі, цитология үшін жаңа 16 түрдің кариотиптік сипаттамасымен. Генетика, 65 (1), 89-107.
- ^ а б Вәли, Ү. (2002). Ескі әлем бүркіттеріндегі митохондриялық псевдо-бақылау аймағы (Aquila тұқымдасы). Молекулалық экология, 11 (10), 2189-2194.
- ^ Lerner, H. R., & Mindell, D. P. (2005). Ядролық және митохондриялық ДНҚ-ға негізделген бүркіттердің, ескі әлемнің қарақұйрықтарының және басқа Accipitridae филогенезі. Молекулалық филогенетика және эволюция, 37 (2), 327-346.
- ^ а б Уотсон, Джефф (2010). Алтын бүркіт. A&C Black. ISBN 978-1-4081-1420-9.
- ^ Stresemann, E., & Amadon, D. (1979). Falconiformes-ге тапсырыс беріңіз. Әлем құстарының тізімі, 1, 271-425.
- ^ Ховард, Р., және Мур, А. (1991). Әлем құстарының толық тізімі (№ 2 ред.). Academic Press Ltd ..
- ^ а б в г. e f ж Кларк, W. S. (1992). "Музей үлгілері мен тірі бүркіттерді бөлу критерийлері бар Дала және Тауан бүркіттерінің таксономиясы" (PDF). 112 (3). Өгіз. B.O.C.: 150–157. Алынған 2019-07-29. Журналға сілтеме жасау қажет
| журнал =(Көмектесіңдер) - ^ Олсон, С.Л. (1994). "Тавни мен Дала Бүркіттерінің бас сүйек остеологиясы Aquila rapax and A. nipalensis" (PDF). 114. Өгіз. B.O.C.: 264–267. Журналға сілтеме жасау қажет
| журнал =(Көмектесіңдер) - ^ Сангстер, Г.; Нокс, А.Г .; Хельбиг, А.Ж .; Паркин, Д.Т. (2002). "Еуропалық құстарға арналған таксономиялық ұсыныстар". Ибис. 144 (1): 153–159. дои:10.1046 / j.0019-1019.2001.00026.x.
- ^ Amadon, D., & Short, L. L. (1976). Түр түріне жақындайтын кіші түрлерді емдеу. Жүйелі зоология, 25 (2), 161-167.
- ^ Brooke, R. K., Grobler, J. H, Irwin, M. P. S., & Steyn, P. (1972). Оңтүстік Африкадағы Aquila nipalensis және A. pomarina (Aves: Accipitridae) көші-қон бүркіттерін зерттеу, басқа ірі рапторлар туралы салыстырмалы жазбалармен. Окк. Нат. Қағаздары Мус. Родезия В5 (2): 61–114.
- ^ Global Raptors
- ^ White, C. M., Olson, P. D., & Kiff, L. F. (1994). Falconidae тұқымдасы (сұңқар мен бүркіт). Әлем құстарының анықтамалығы, 2: 52-214.
- ^ а б Kemp, AC және G. M. Kirwan (2020). Тауан бүркіті (Aquila rapax), 1.0 нұсқасы. Әлем құстарында (Дж. Дель Хойо, А. Эллиотт, Дж. Саргатал, Д. А. Кристи және Э. де Хуана, редакторлар). Орнитологияның Корнелл зертханасы, Итака, Нью-Йорк, АҚШ.
- ^ а б в г. e f Ash, C. P., & Atkins, J. D. (2009). Эфиопия мен Эритрея құстары: таралу атласы. A&C Black.
- ^ а б Мусиндо, П.Т. (2012). Оңтүстік Африка жыртқыш құстарындағы жыртқыш типіне қатысты шоттар мен тырнақтардағы морфологиялық өзгеріс (Falconiformes және Strigiformes бұйрықтары).
- ^ а б Clouet, M., Barrau, C., & Goar, J. L. (2000). Эфиопиялық Balé Highlands күндізгі Афро-альпілік рапторлар қауымдастығы. Түйеқұс, 71 (3-4), 380-384.
- ^ Кук, W. E. (2012). Құстардың шөлді жыртқыштары. Springer Science & Business Media.
- ^ Аль-Шейхли, О.Ф., & Аль-Азави, Дж. (2019). Оңтүстік Ирактағы Месопотамия батпақтарындағы жыртқыш құстар (жыртқыштар). Ирак табиғи тарихының мұражайы хабаршысы (P-ISSN: 1017-8678, E-ISSN: 2311-9799), 15 (4), 381-402.
- ^ Crochet, PA, Raty, L., De Smet, G., Anderson, B., Barthel, PH, Collinson, JM, Dubois PJ, Helbig, AJ, Jiguet, F., Jirle, E., Knox, AG, Le Maréchal, P., Parkin, DT, Pons, J.-M., Roselaar CS, Svensson, L., van Loon, AJ, and Yésou P. (2010). AERC TAC таксономиялық ұсынымдары. Шілде 2010. Диспансер: http: // www. aerc. eu / tac. HTML.
- ^ Sibley, C. G., & Monroe, B. L. (1990). Әлем құстарының таралуы және таксономиясы. Йель университетінің баспасы.
- ^ а б в г. e f ж сағ Али, С., & Рипли, С., Д. (1980). Бангладеш, Непал, Бутан және Шри-Ланка сияқты Үндістан мен Пәкістан құстарының анықтамалығы.. Оксфорд университетінің баспасы.
- ^ а б Бейкер, E. C. (1928). Цейлон мен Бирманы қоса алғанда, Британдық Үндістан фаунасы. Тейлор және Фрэнсис.
- ^ а б в Әли, Сәлім (1993). Үнді құстарының кітабы. Бомбей: Бомбей табиғи тарих қоғамы. ISBN 0-19-563731-3.
- ^ а б в г. Ширихай, Х. (1994). Тауанды бүркіттің Израильдегі дала қыранынан бөлінуі. Британдық құстар, 87, 396-397.
- ^ Snelling, J. C. (1969). Кейбір мәліметтер Оңтүстік Африка Республикасы, Крюгер ұлттық паркіндегі ірі рапторларды белгілеуден алынған. Түйеқұс, 40 (S1), 415-427.
- ^ а б CRC құс дене массасының анықтамалығы, 2-шығарылым Джон Б. Даннинг кіші (редактор). CRC Press (2008), ISBN 978-1-4200-6444-5.
- ^ Borrow, N., & Demey, R. (2013). Гана құстарына арналған далалық нұсқаулық. Bloomsbury Publishing.
- ^ Ван Перло, Б. (1999). Африканың оңтүстігіндегі құстар. Harpercollins Pub Limited.
- ^ Тауан бүркіті (Акила рапаксы), ARKive, 2011, мұрағатталған түпнұсқа 2018-02-01
- ^ Мендельсон, Дж. М., Кемп, А.С., Биггз, Х.С., Биггс, Р., & Браун, Дж. (1989). Африка рапторларының 66 түрінің қанаттарының аймақтары, қанаттарының жүктемелері және қанаттарының аралықтары. Түйеқұс, 60 (1), 35-42.
- ^ а б в г. e Андерсон, Дж., & Хорвиц, Р. Дж. (1979). Бүркіттер мен олардың құс бәсекелестері арасындағы бәсекелестік өзара іс-қимыл. Ибис, 121 (4), 505-509.
- ^ а б в г. e f ж сағ Хоккей, П.Р., Дин, В.Р.Ж. & Райан, П.Г. (2005). Робертс - Африканың оңтүстігіндегі құстар, VII-ші басылым. Джон Фоулкердің құс кітап қорының қамқоршылары, Кейптаун.
- ^ а б в г. e f ж сағ мен j к л м n o б q р с т сен v w х ж Сминк, C. (1974). Шығыс Африканың кейбір жыртқыш құстарын салыстырмалы-экологиялық зерттеулер. Ардея 62 (1-2): 1-97.
- ^ Кемп, А., & Кемп, М. (2006). Сасол жыртқыш құстары; Жаңа басылым. Струк.
- ^ а б Форсман, Д. (1999). Еуропа мен Таяу Шығыстың рапторлары: өрісті сәйкестендіру бойынша анықтамалық. Лондон: T & AD Poyser.
- ^ Янковиц, М. (1976). Таван немесе дала қыраны. Бокмакиери, 28 (3), 64-65.
- ^ а б в г. Дэвидсон, И. (1978). Африканың оңтүстік рапторларын ұшуды анықтау. Бокмакиери, 30: 43-48.
- ^ а б в г. e f ж сағ мен j к л м n o б q р с т сен v w х ж з аа аб ак жарнама ае аф Стейн, П. (1973). «Тауан бүркітіндегі бақылаулар". Түйеқұс. 44 (1): 1–22. дои:10.1080/00306525.1973.9632611.
- ^ Дэвисон, Б. (1998). Зимбабвенің оңтүстік-шығысындағы мекендейтін рэпторлар қауымдастығы (Докторлық диссертация, Родос университеті).
- ^ Isenmann P. & Moali A. (2000). Алжир құстары. Société d ‟Ornithologiques de France, Париж.
- ^ Николаус, Г. (1987). Судан құстарының таралу атласы мекендеу орны мен жағдайы туралы жазбалармен. Zoologisches Forschungsinstitut und музейі Александр Кениг.
- ^ а б Смит, К.Д. (1957). Эритрея құстарының түсіндірме тізімі. Ибис, 99 (2), 307-337.
- ^ Ash, J. S., & Miskell, J. E. (1998). Сомали құстары. Pica Press.
- ^ а б Тевенот, М., Вернон, Р., және Берджер, П. (2003). Марокконың құстары: түсіндірме тізімі (№ 20). Британдық орнитологтар одағы.
- ^ Исенманн, П. (2005). Тунис құстары. Société d'études ornithologiques de France, Muséum ұлттық d'histoire naturelle.
- ^ Borrow, N., & Demey, R. (2001). Батыс Африканың құстарына арналған нұсқаулық. Принстон, Ндж.
- ^ Гор, М.Э. Дж. (1990). Гамбия құстары. Бақылау тізімі 3. Британдық орнитологтар одағы, Тринг.
- ^ а б в Тиоллей, Дж. М. (1985). Кот-д'Ивуардың құстары: жағдайы және таралуы. Батыс Африка орнитологиялық қоғамы.
- ^ Greig-Smith, P. W. (1976). Моле ұлттық паркінің авифаунасының құрамы мен тіршілік ету ортасы, Гана. Өгіз. Нигериялық орн. Soc, 12 (42), 45-66.
- ^ а б Dowsett, R. J., Dowsett-Lemaire, F., & Hester, A. (2008). Гана авифауна: толықтырулар мен түзетулер. Өгіз. Афр. Құстар клубы, 15, 191-200.
- ^ а б Cheke, R. A., & Walsh, J. F. (1996). Того құстары: түсіндірме парағы (№ 14). Британдық орнитологтар одағы.
- ^ Элгуд, Дж. Х., Хейгам, Дж.Б., Мур, А.М., Нейсон, А.М., Шарланд, Р.Э. & Скиннер, Н. Дж. (1994). Нигерия құстары: түсіндірме тізімі. Британдық орнитологтар одағы.
- ^ Carswell, M., Pomeroy, D., Reynolds, J., & Tushabe, H. (2005). Угандадағы құс атласы. Британдық орнитологтар клубы және британдық орнитологтар одағы.
- ^ Dowsett-Lemaire, F., & Dowsett, R. J. (2006). Малави құстары: атлас және анықтамалық. Tauraco & Aves.
- ^ Стивенсон, Т., және Фаншоу, Дж. (2002). Шығыс Африка құстарына арналған далалық нұсқаулық. T & AD Poyser.
- ^ а б в г. Dowsett, J. J., Aspinwall, D. R., & Dowsett-Lemaire, F. (2008). Замбия құстары: атлас және анықтамалық. Tauraco Press.
- ^ Дин, W. R. J. (2000). Анголаның құстары: түсіндірмелі бақылау парағы. Британдық орнитологтар одағы.
- ^ da Rosa Pinto, A. A. (1983). Ornitologia de Angola (1-том). Instituto de Investigação Científica Tropical.
- ^ Паркер, В. (2005). Мозамбиктің орталық құстарының атласы. Жойылу қаупі төнген жабайы табиғатқа сенім және құстардың демографиялық бөлімі.
- ^ а б в г. e Ирвин, M. P. S. (1981). Зимбабве құстары. Quest Pub ..
- ^ а б Penry, H. (1994). Ботсвананың құс атласы. Квазулу университеті Natal Press.
- ^ Хэнкок, П., Мюллер, М., және Тайлер, Дж. (2007). Рамазан Окаванго атырауындағы құстардың тізімдемесі. Бабблер, 49, 3-29.
- ^ Барнс, К.Н. (Ред.) (2000). Эскомның Оңтүстік Африка, Лесото және Свазиленд құстарының қызыл кітабы. BirdLife Оңтүстік Африка.
- ^ Boshoff, A. F., & Vernon, C. J. (1980). Кейп провинциясындағы кейбір бүркіттердің таралуы және мәртебесі. Кейп провинциясы мұражайларының жылнамалары (табиғи тарих), 13 (9), 107-132.
- ^ Monadjem, A., & Rasmussen, M. W. (2008). Свазилендтегі бүркіттердің, таңдалған сұңқарлар мен үкілердің ұясының таралуы және консервация күйі. Габар, 19, 1-22.
- ^ Дженнингс, М.С. (1991). Сауд Арабиясының өзгеріп жатқан авифауна. Сауд Арабиясы Табиғат тарихы қоғамының журналы, 3, 16-28.
- ^ Мартинс, Р.П. және Портер, Р.Ф. (Ред.). (1996). Оңтүстік Йемен және Сокотра: OSME зерттеуінің 1993 жылғы көктемдегі есебі. Таяу Шығыстың орнитологиялық қоғамы.
- ^ Shirihai, H., Dovrat, E., Christie, D. A., & Harris, A. (1996). Израиль құстары. Лондон: Academic Press.
- ^ Эриксен, Дж., Саргеант, Д.Е. & Виктор, Р. 2003 ж. Оман құстарының тізімі (6-шы шығарылым). Экологиялық зерттеулер мен зерттеулер орталығы, Сұлтан Кабус университеті, Маскат, Оман
- ^ а б в г. e f Расмуссен, П.С. және Андертон, Дж.С. (2005). Оңтүстік Азияның құстары: Рипли гид. Вашингтон, ДС.
- ^ Скотт, Д.А, және Адхами, А. (2006). Иран құстарының жаңартылған бақылау тізімі. Подокес, 1 (1/2), 1-16.
- ^ Roth, T., Ayé, R., Burri, R., & Schweizer, M. (2005). 2001 жылдың ақпан-наурыз айларында Ираннан құстардың бақылаулары, соның ішінде Таяу Шығыс үшін жаңа түр. Сандгроуз, 27: 63-68.
- ^ Inskipp, C. & Inskipp, T. (1991). Непал құстарына арналған нұсқаулық. Екінші басылым. Кристофер Хельм, Лондон.
- ^ Дональд, Х. (1919). Месопотамияның кейбір жыртқыш құстары. Бомбей табиғи тарих қоғамының журналы, 26: 845-846.
- ^ а б Duckworth, J. W., Inskipp, T. P., Pasquet, E., Rasmussen, P. C., Rays, N. H., Robson, C. R., & Russell, D. G. D. (2008). Tawny Eagle Aquila рапаксының Оңтүстік-Шығыс Азиядағы мәртебесін қайта бағалау.
- ^ Варакагода, Д. (2015). Шри-Ланка құстарына арналған фотографиялық нұсқаулық. A & C Black Limited.
- ^ Фрай, Х. (1966). Солтүстік Гвинея саваннасында, Нигерияда құстардың экологиялық таралуы. Түйеқұс, 37 (1): 335-356.
- ^ а б Хастлер, К., және Хауэллс, В.В. (1990). Бастапқы өндірістің Хвандж ұлттық паркіндегі рэпторлар қауымына әсері, Зимбабве. Тропикалық экология журналы, 6 (3), 343-354.
- ^ а б в г. e Херхолдт Дж .; Кемп, А .; Du Plessis, D. (1996). «Калахари Джемсбок ұлттық паркіндегі Бейтел мен Тауан бүркітінің өсу мәртебесі мен экологиясының аспектілері, Оңтүстік Африка". Түйеқұс. 67 (3–4): 126–137. дои:10.1080/00306525.1996.9639697.
- ^ Malan, G., & Howells, B. (2009). Ағаштардың ұясын зерттеу. Рапторлық зерттеу және бақылау: Африкалық жыртқыш құстарға арналған далалық нұсқаулық, 42.
- ^ а б Тиоллей, Дж. М. (1992). Батыс Африкадағы эфиопиялық рапторлардың маусымдық қоныс аударуының заңдылықтары мен экологиясы. 7-Африка орнитологиялық конгресінде, Найроби (28-том).
- ^ Snelling, J. C. (1970). Кейбір мәліметтер Оңтүстік Африка Республикасы, Крюгер ұлттық паркіндегі ірі рапторларды белгілеуден алынған. Түйеқұстың қосымшасы, 8: 415-427.
- ^ Ławicki, Ł., Jiguet, F., El Din, S. B., van den Berg, A. B., Corso, A., Crochet, P. A., Hoath, R., Schweizer, M. & Waheed, A. (2019). Египет орнитологиялық сирек кездесетін комитетінің алтыншы есебі.
- ^ Арнольд, П.К (1965). Израиль құстары. Shalit Publishers.
- ^ Кинир, Н.Б (1931). Орталық Арабияның кейбір құстарында. Ибис, 73 (4), 698-701.
- ^ Джахан, И., Томпсон, П.М., Пол, Е., Султана, Н., Рой, К., & Маллик, А. (2017). Бхавал ұлттық паркі құстары, Бангладеш. Форктаил, (33), 88-91.
- ^ Oatley, TB, Oschadleus, HD., Navarro, RA. & Underhill, Л.Г. (1998). Африканың оңтүстігінде жыртқыш құстардың сақиналық қалпына келтірілуіне шолу: 1948-1998 жж. Жойылып кету қаупі бар жабайы табиғат қоры, Йоханнесбург.
- ^ Wichmann, M.C., Groeneveld, J., Jeltsch, F., & Grimm, V. (2005). Климаттың өзгеруіне рапорларға әсерді мінез-құлыққа бейімдеу арқылы азайту: экологиялық буферлік механизмдер. Ғаламдық және планеталық өзгерістер, 47 (2-4), 273-281.
- ^ Шакил, М. (2004). Акила Рапакс. In: Кашмир қырандары. Өзін-өзі жариялады.
- ^ Дармакумарсинхжи, Р.С. (1949). Жыртқыш құстар серік ретінде. Авикультуралық журнал, 55: 81-85.
- ^ Эллис, Д.Х., Беднарз, Дж.С., Смит, Д.Г. және Флемминг, С.П. (1993). Рапториалды құстардағы әлеуметтік қоректену сыныптары: Жоғары дамыған кооперативтік аңшылық көптеген рапторлар үшін маңызды болуы мүмкін. Биология, 43 (1), 14-20.
- ^ Барбур, Д. (1975). Бүркіттер аң аулау үшін ай сәулесін қолдана ма? Honeyguide, 82: 37.
- ^ а б в г. e f ж сағ мен j к л м Осборн, Т. (1982). «Оңтүстік Замбиядағы Тауан бүркітіне бақылау". Түйеқұс. 53 (2): 107–111. дои:10.1080/00306525.1982.9634734.
- ^ Vernon, C. J. (1979). Жыртқыш жеті Тауан бүркіт ұясынан қалады. Медовый гид, 100: 22-24.
- ^ а б в г. e f ж сағ мен Тарботон, В.Р. & Аллан, Д.Г. (1984). Трансваальда жыртқыш құстардың жағдайы мен сақталуы. Трансвааль мұражайы Монография № 3. Претория.
- ^ Boshoff, A. F., Rous, R. C., & Vernon, C. J. (1981). Кейп провинциясындағы Колесберг ауданындағы аңшы бүркіттің жемтігі. Түйеқұс, 52 (3), 187-188.
- ^ Мюллер, Дж. П. 1977 ж. Arvicanthis abyssinicus in Der Grassteppe des Semien Mountain National Park (Athiopien) популяциясы. З.Саугетерк, 42 (3): 145-172.
- ^ а б в Ван Сомерен, В.Г.Л (1956). Құстармен күндер: кейбір Шығыс Африка түрлерінің әдеттерін зерттеу (38-том). Чикаго табиғи тарих мұражайы.
- ^ а б в г. Дармакуарсинхжи, К.С. (1955). Саураштра құстары. Дил Бахар.
- ^ а б Fitzwater, W. D., & Prakash, I. (1969). Мерионес хуррианае Джердон үнді шөлінің гербінің шұңқырларына, мінез-құлқына және үй аймағына бақылау.. Сүтқоректілер, 33 (4), 598-606
- ^ а б Prakash, I., & Ghosh, P. K. (Eds.). (2012). Шөл ортасындағы кеміргіштер (28-том). Springer Science & Business Media.
- ^ Джибан, Дж. (1978). Бурунди Республикасының суармалы егістігінде мастомисматикалық егеуқұйрықты, Mastomys natalensis (А.Смит) ауруын бақылау. 8-ші омыртқалы зиянкестер конференциясының материалдарында (1978) (20-бет).
- ^ а б Кингдон, Дж., Хаппольд, Д., Бутинский, Т., Гофман, М., Хапполд, М., және Калина, Дж. (2013). Африканың сүтқоректілері. A&C Black.
- ^ Mbise, F. P., Fredriksen, KE, Fyumagwa, R. D., Holmern, T., Jackson, C. R., Fossøy, F., and Røskaft, E. (2017). Серенгетидегі адамның қатысуынан гиракстардың пайдасы бар ма? Африканың экология журналы, 55 (4), 672-678.
- ^ Дин, В.Р. Дж., Хантли, М.А., Хантли, Дж., & Вернон, Дж. Дж. (1988). Анголаның кейбір құстарына арналған ескертпелер. Novitates, Дурбан мұражайы, 14 (4), 43-92.
- ^ Бейнон, П., & Раса, О.А. (1989). Ергежейлі моңғолдардың тілді ескертуге арналған дауыстары күрделі ақпаратты жібере ме? Оңтүстік Африка ғылымдар журналы, 85 (7): 447-450.
- ^ Manser, M. B., Seyfarth, R. M., & Cheey, D. L. (2002). Суратикалық дабыл қоңырау жыртқыш класы мен жеделдігін білдіреді. Когнитивті ғылымдардағы тенденциялар, 6 (2), 55-57.
- ^ Тарботон, В.Р., Пикфорд, П., & Пикфорд, Б. (1990). Африка жыртқыш құстары. Корнелл университетінің баспасы.
- ^ Malcom, J. R. (1986). Құлақты түлкілердің әлеуметтік экологиясы (Otocyon megalotis). Зоология журналы, 208 (3), 457-469.
- ^ Mercier, S., Neumann, C., van de Waal, E., Chollet, E., de Bellefon, J. M., & Zuberbühler, K. (2017). Вервет маймылдары ересек еркектерге қауіпті жағдайларда сәлем береді. Жануарлардың мінез-құлқы, 132, 229-245.
- ^ Перейра, М.Э. (1988). Жас пен саваннаның бабундарындағы, Papio cynocephalus cynocephalus ішіндегі интервалдық тәртіпке жас пен жыныстың әсері. Жануарлардың мінез-құлқы, 36 (1), 184-204.
- ^ Пахад, Г. (2010). Суйкербсранд қорығының чакма бабундарындағы әлеуметтік мінез-құлық және өсімдіктерді рейдерлік (Докторлық диссертация, Витватерсранд университеті).
- ^ Zinner, D., & Peláez, F. (1999). Верреаның бүркіттері (Aquila verreauxi) Эритреядағы хамадрия бабуондарының (Papio hamadryas hamadryas) ықтимал жыртқыштары ретінде. Американдық Приматология журналы: Американдық Приматологтар Қоғамының ресми журналы, 47 (1), 61-66.
- ^ Робертс, Б.А. (2014). Аналықты сынау: Томсонның жейреніндегі аналық жүріс-тұрыс және антипредатортактика (Gazella thomsonii) (Докторлық диссертация, Принстон университеті).
- ^ Dixon, J. E. W. (1968). Ірі рапторлардың жемтігі. Түйеқұс, 39 (3), 203-204.
- ^ Douthwaite, R. J. (1978). Замбиядағы Кафуе пәтерінде қаздар мен қызыл ысталған кот, 1970–1974 жж. Африка журналы Экология журналы, 16 (1), 29-47.
- ^ Хоссейни-Моосави, С.М., Бехрузи-Рад, Б., Каримпур, Р., & Насаб, С.М.А. (2013). Иранның оңтүстік-батысында, Хузестан провинциясында жағалы көгершін (Streptopelia decaocto) өсіру биологиясы. Беркут, 22 (1), 51-54.
- ^ Combreau, O., & Smith, T. R. (1998). Сауд Арабиясында губараның көкірекшелерін енгізу кезінде босату техникасы мен жыртқыштық. Биологиялық консервация, 84 (2), 147-155.
- ^ Руно, М. (2000). Қызыл бүркітпен тамақтанатын Таван Бүркіт. Медовый гид, 46:24.
- ^ Уолтерс, Дж. Р. (1990). Лепвингтердің жыртқышқа қарсы мінез-құлқы: дискриминациялық қабілеттердің далалық дәлелі. Уилсон бюллетені, 49-70.
- ^ Робинсон, В. (2015). Шығыс африкалық сода көлдерінің экологиясы: кішігірім фламинго (кішігірім Phoeniconaias) үшін тамақтану тәртібі (Ph.D. Диссертация). Лестер, Англия: Лестер университеті.
- ^ Джонсон, А., & Сезилли, Ф. (2009). Үлкен қоқиқаз. A&C Black.
- ^ Лихтенберг, E. M., & Hallager, S. (2008). Kori bustard үшін жиі байқалатын мінез-құлықтың сипаттамасы (Ardeotis kori). Этология журналы, 26 (1), 17-34.
- ^ Джексон, Х.Д., & Слотов, Р. (2002). Афротропикалық түнгі уақыттағы өлім-жітімге шолу, негізінен адам өлтіреді. Түйеқұс, 73 (3-4), 147-161.
- ^ Раджу Касамбе, Г. (2010). Көк құйрықты аралар - Мумбай мен оның айналасындағы «қыста қоныс аударушылар». Құстарды бақылаушыларға арналған ақпараттық бюллетень, 50 (3), 33.
- ^ Брожерова, Дж. (2013). Referenční vokalizace papouška žako kongo (Psittacus erithacus).
- ^ Дин, W. R. J. (2004). Көшпенді шөлді құстар: 32 үстелмен. Springer Science & Business Media.
- ^ ван Херден Дж. (1977). Quelea жыртқыштары. Бокмакиери, 29: 45.
- ^ Vernon, C. J. (1967). KW Greenhow журналдарынан алынған кейбір ескертулер. Түйеқұс, 38 (1), 48-49.
- ^ Парри-Джонс, Дж. (1997). Бүркіт. Dorling Kindersley Ltd.
- ^ а б в Браун, Л.Х. (1952). Кения колониясы - Эмбу ауданының ірі жыртқыш құстарының биологиясы туралы. Ибис, 94 (4), 577-620.
- ^ Ramesh, M., & Sankaran, R. (2013). Тар аймағындағы Үндістанның тікенді кесірткесі Uromastyx hardwickii туралы табиғи тарихты бақылау. Раджастханның Фаунальдық мұрасында, Үндістан (295-310 беттер). Спрингер, Нью-Йорк, Нью-Йорк.
- ^ Байндур, А. (2009). Рапторлар мен агамидтер. Үнді құстары, 5 (1), 11-13.
- ^ Хауэллс, В.В. (1978). Жыртқыш балықтар ірі жыртқыш құстардың қорек көзі ретінде. Медовый гид, 96: 18.
- ^ Dial, K. P., & Vaughan, T. A. (1987). Кениядағы алат термиттеріне арналған оппортунистік жыртқыштық. Биотропика, 19 (2), 185-187.
- ^ Bussia, E., & Wijers, M. (2013). Жемді ашуландыру: термиттер тобында 50-ден астам рапторлар. Биоалуантүрлілікті бақылау, 11-18.
- ^ Anene, C., & Vajime, C. G. (1990). Нигериядағы Oedaleus senegalensis Krauss (Orthoptera: Acrididae) паразиттері, паразитоидтары және жыртқыштары.. Халықаралық тропикалық жәндіктер туралы журнал, 11 (1), 27-34.
- ^ ван Джаарсвельд, Дж. (1987). Тауан бүркіт пен аққұба пілдің тезегінен бірге тамақтанады. Габар, 2:26.
- ^ а б в г. Kemp, A. C., & Kemp, M. I. (1975). Крюгер ұлттық паркіндегі ақ сүйекті сығандар сығандар африканусындағы бақылаулар, басқа құс тазалаушылар туралы жазбалар. Коедое, 18 (1), 51-68.
- ^ Mulualem, G., Tesfahunage, W., & Ayalew, S. (2015). Эфиопияның шығыс аймағында орналасқан авифаунаның жергілікті қарақұйрық мейрамханасында көптігі мен белсенділігі. Халықаралық құс және жабайы табиғат биология журналы, 1 (1).
- ^ а б в г. e f ж Mundy, PJ (1982). Африка оңтүстік қарақұйрықтарының салыстырмалы биологиясы. Йоханнесбург. Vulture Study тобы.
- ^ а б Dean, W. R. J., & MacDonald, I. A. W. (1981). A review of African birds feeding in association with mammals. Ostrich, 52(3), 135-155.
- ^ а б в г. Ogada, D.L.; Монаджем, А .; McNally, L.; Kane, A.; Jackson, A.L. (2014). "Vultures acquire information on carcass location from scavenging eagles". Корольдік қоғамның еңбектері B: Биологиялық ғылымдар. 281 (1793): 20141072. дои:10.1098/rspb.2014.1072. PMC 4173674. PMID 25209935.
- ^ а б в Brown, C. J. (1982). The behaviour of a Tawny Eagle at carrion. Madoqua, 14 (1): 95-97.
- ^ а б в г. e f ж сағ мен Kendall, C. J. (2013). Alternative strategies in avian scavengers: how subordinate species foil the despotic distribution. Behavioral Ecology and Sociobiology, 67(3), 383-393.
- ^ Plaskett, G. (1972). Tawny Eagle/vulture behaviour. Witwaterstrand Bird Club News Sheet, 80: 11.
- ^ а б в Houston, D.C. (1974). The role of griffon vultures Gyps africanus and Gys rueppellii as scavengers. J. of Zoo, London. 171: 35-46.
- ^ Pomeroy, D. E. (1975). Birds as scavengers of refuse in Uganda. Ibis, 117(1), 69-81.
- ^ Mesfin, K. (2014). Diversity and Abundance of Birds in Dbla Church Forest, Eastern Tigray, North Ethiopia. Journal of Zoology Studies, 1(5), 1-8.
- ^ Samson, A., & Ramakrishnan, B. (2017). Scavenging Mode of Vertebrate Scavengers on Domestic Buffalos Bubalus bubalis (Linnaeus, 1785) Killed by Tiger Panthera tigris and Natural Deaths in Southern India. Journal homepage: www. wesca. net, 12(1).
- ^ Lewis, A.D. (1990). Tawny Eagle Aquila rapax following a foraging pack of Hunting Dogs Lycaon pictus. Scopus, 14: 19-20.
- ^ Brockmann, H. J., & Barnard, C. J. (1979). Kleptoparasitism in birds. Animal behaviour, 27, 487-514.
- ^ Olsen, K. M. (2013). Skuas and jaegers. A&C Black.
- ^ Corre, M. L., & Jouventin, P. (1997). Kleptoparasitism in tropical seabirds: vulnerability and avoidance responses of a host species, the Red-footed Booby. The Condor, 99(1), 162-168.
- ^ Haynes, J.R. (1981). Tawny Eagle pirates food from a Dark Chanting-goshawk. Honeyguide, 50.
- ^ Leonardi, G. (2015). The lanner falcon. Italy: Giovanni Leonardi.
- ^ Kemp, A. C., & Kemp, M. I. (1980). The biology of the southern ground hornbill Bucorvus leadbeateri (Vigors)(Aves: Bucerotidae). Annals of the Transvaal Museum, 32(4), 65-100.
- ^ Kemp, M. I., & Kemp, A. C. (1978). Bucorvus and Sagittarius: Two modes of terrestrial predation. In: Proceedings of the Symposium on African Predatory Birds, Northern Transvaal Ornithological Society, Pretoria.
- ^ а б Watson, R. T., & Watson, C. (1987). Interspecific piracy between Tawny Eagles and Bateleurs: how common is it. Gabar, 2, 9-11.
- ^ Krueger, O. (1997). Population density and intra‐and interspecific competition of the African Fish Eagle Haliæeetus vocifer in Kyambura Game Reserve, southwest Uganda. Ibis, 139(1), 19-24.
- ^ Meyburg, B. U., & Kirwan, G. M. (2018). Eastern Imperial Eagle (Aquila heliaca). Handbook of the Birds of the World Alive (J. del Hoyo, A. Elliott, J. Sargatal, DA Christie, and E. de Juana, Editors). Lynx Edicions, Барселона, Испания.
- ^ Clouet, M., Barrau, C. & Goar, J.L. (1999). The Golden Eagle (Aquila chrysaetos) in the Bale Mountains, Ethiopia. Raptor зерттеу қоры, 33 (2): 102-109.
- ^ Sillero-Zubiri, C., & Gottelli, D. (1995). Diet and feeding behavior of Ethiopian wolves (Canis simensis). Journal of Mammalogy, 76(2), 531-541.
- ^ Ali, H. (2008). Behaviour and ecology of the House Crow (Corvus splendens) in Islamabad-Rawalpindi and Adjoining areas. University of Agriculture, Faisalabad, Pakistan.
- ^ Watson, R. T. (1986). The ecology, biology and population dynamics of the Bateleur Eagle (Terathopius ecaudatus) (Doctoral dissertation, RT Watson).
- ^ Bildstein, K. L., & Parry-Jones, J. (2011). The eagle watchers: Observing and conserving raptors around the world. Корнелл университетінің баспасы.
- ^ Machange, R. W., Jenkins, A. R., & Navarro, R. A. (2005). Eagles as indicators of ecosystem health: Is the distribution of Martial Eagle nests in the Karoo, South Africa, influenced by variations in land-use and rangeland quality? Journal of arid environments, 63(1), 223-243.
- ^ а б в Engel, J. I. (2011). Possible predation of a Pygmy Falcon by a Tawny Eagle in Namibia. Biodiversity Observations, 34-35.
- ^ Van Nieuwenhuyse, D., Genot, J. & Johnson, D. (2008). The Little Owl: Conservation, Ecology and Behavior of Athene noctua. Кембридж, Нью-Йорк: Кембридж университетінің баспасы.
- ^ Begg, C. M., Begg, K. S., Du Toit, J. T., & Mills, M. G. L. (2003). Sexual and seasonal variation in the diet and foraging behaviour of a sexually dimorphic carnivore, the honey badger (Mellivora capensis). Journal of Zoology, 260(3), 301-316.
- ^ Murn, C., Betchley, P., & Robert, C. (2009). Talon-locking and cartwheeling as a prelude to copulation in Tawny Eagles Aquila rapax. Gabar, 20, 12-14.
- ^ Borello, W. D., & Borello, R. M. (2004). Two incidents of talon-grappling and cartwheeling in the Tawny Eagle Aquila rapax. Ostrich-Journal of African Ornithology, 75(4), 320-321.
- ^ Simmons, R. E., & Mendelsohn, J. M. (1993). A critical review of cartwheeling flights of raptors. Ostrich, 64(1), 13-24.
- ^ а б Vincent, A. W. (1945). On the breeding habits of some African birds. Ibis, 87(3), 345-365.
- ^ Anderson A. (1871). Notes on the raptorial birds of India. Journal of Zoology, 44: 675-690.
- ^ а б в г. e f Дженкинс, А.Р .; De Goede, K.H.; Lovelater, S.; Diamond, M. (2013). "Brokering a settlement between eagles and industry: sustainable management of large raptors nesting on power infrastructure" (PDF). Халықаралық құстарды қорғау. 23 (2): 232–246. дои:10.1017/S0959270913000208. Алынған 2019-07-29.
- ^ Maclean, G. L. (1969). The breeding seasons of birds in the south-western Kalahari. Ostrich, 40(1): 179-192.
- ^ Tarboton, W. R., Kemp, M. I., & Kemp, A. C. (1987). Birds of the Transvaal. Трансвааль мұражайы.
- ^ Kendall, C.J.; Rubenstein, D.L.; Slater, P.L.; Монаджем, А. (2017). "An assessment of tree availability as a possible cause of population declines in scavenging raptors". Құс биологиясының журналы. 49 (1): 001–008. дои:10.1111/jav.01497. Алынған 2019-07-29.
- ^ de Balsac, H. H., & Mayaud, N. (1962). Les oiseaux du nord-ouest de l'Afrique: distribution géographique, écologie, migrations, reproduction (Vol. 10). P. Lechevalier.
- ^ Bannerman, D. A. (1951) The birds of tropical West Africa. Crown Agents, London.
- ^ Bannerman, D. A. (1953). The Birds of West and Equatorial Africa. Эдинбург және Лондон.
- ^ McGowan, G. (1992). Tawny Eagle pair raise two chicks. Babbler, 23: 50.
- ^ а б Hustler, K., & Howells, W. W. (1986). A population study of tawny eagles in the Hwange National park, Zimbabwe. Ostrich, 57(2), 101-106.
- ^ Flower, S. S. (1923). List of Birds of Prey 1898-1923: With Notes on Their Longevity. Government Press.
- ^ Underhill, L.; Brooks, M. (2014). "Preliminary summary of changes in bird distributions between the first and second southern African Bird Atlas projects (SABAP AND SABAP2)". Орнитологиялық бақылаулар. 5: 258–293. Алынған 2019-07-29 – via University of Cape Town Libraries.
- ^ а б в г. Brown, C. J. (1991). Declining Martial Polemaetus bellicosus and Tawny Aquila rapax Eagle populations and causes of mortality on farmlands in central Namibia. Biological conservation, 56(1), 49-62.
- ^ Herholdt, J. J. (1998). Survival threats and conservation management of raptors in the Kgalagadi Transfrontier Park. Transactions of the Royal Society of South Africa, 53(2), 201-218.
- ^ Thiollay, J. (2006). «The decline of raptors in West Africa : long-term assessment and the role of protected areas". IBIS. 148 (2): 240–254. дои:10.1111/j.1474-919X.2006.00531.x.
- ^ "Over 500 Rare Vultures Die After Eating Poisoned Elephants In Botswana". France-Press агенттігі. NDTV. 2019-06-21. Алынған 2019-06-28.
- ^ а б Хурворт, Элла (2019-06-24). "More than 500 endangered vultures die after eating poisoned elephant carcasses". CNN. Алынған 2019-06-28.
- ^ Солли, Мейлан (2019-06-24). «Ботсванада браконьерлердің улары 530 жойылып бара жатқан қарақұйрықты өлтірді». Смитсониан. Алынған 2019-06-28.
- ^ Нгуоу, Борис (2019-06-27). «Ботсвана: 500-ден астам қарақұйрық жаппай уланудан кейін өлі табылды». Африка21. Алынған 2019-06-28.
- ^ Loftie-Eaton, M. (2014). Geographic range dynamics of South Africa’s bird species. South Africa: Department of Biological Sciences, University of Cape Town.
- ^ Virani, M. Z., Kendall, C., Njoroge, P., & Thomsett, S. (2011). Major declines in the abundance of vultures and other scavenging raptors in and around the Masai Mara ecosystem, Kenya. Biological Conservation, 144(2), 746-752.
- ^ Dutta, T. K., Sudhan, N. A., & Azmi, S. (2008). PCR detection of Pasteurella multocida in eagle PCR based detection, characterization and antibiogram of Pasteurella multocida from wild eagle (Aquila rapax) in Jammu and Kashmir. The Indian Journal of Animal Sciences, 78(12).
- ^ Wichmann, M. C., Jeltsch, F., Dean, W. R. J., Moloney, K. A., & Wissel, C. (2003). Implication of climate change for the persistence of raptors in arid savanna. Oikos, 102(1), 186-202.
- ^ Van Rooyen, C. S. (2000). Raptor mortality on power lines in South Africa. In Raptors at risk: proceedings of the 5th world conference on birds of prey and owls. Hancock House Publishers and the World Working Group on Birds of Prey and Owls, Blaine, Washington (pp. 739-750).
- ^ Anderson, M. D., Maritz, A. W., & Oosthuysen, E. (1999). Raptors drowning in farm reservoirs in South Africa. Ostrich, 70(2), 139-144.
- ^ Pande, S., Padhye, A., Deshpande, P., Ponkshe, A., Pandit, P., Pawashe, A., Pednekar , S., Pandit, R. & Deshpande, P. (2013). CEPF Western Ghats Special Series: Avian collision threat assessment at Bhambarwadi Wind Farm Plateau in northern Western Ghats, India. Journal of Threatened Taxa, 5(1), 3504-3515.
- ^ Crick, H. Q., & Sparks, T. H. (1999). Climate change related to egg-laying trends. Nature, 399(6735), 423-423.
- ^ Aumann, T. (2001). The structure of raptor assemblages in riparian environments in the south-west of the Northern Territory, Australia. Emu, 101(4), 293-304.
- ^ Post, E., Peterson, R. O., Stenseth, N. C. & McLaren, B. E. (1999). Ecosystem consequences of wolf behavioural responses to climate. Nature, 401: 905–907.
- ^ Moss, R., Oswald, J., & Baines, D. (2001). Climate change and breeding success: decline of the capercaillie in Scotland. Journal of Animal Ecology, 47-61.
Сыртқы сілтемелер
- Tawny eagle - Species text in Оңтүстік Африка құстарының атласы
- Тауан бүркіт кезінде Жануарлардың алуан түрлілігі