Қызыл импортталған өрт құмырсқасы - Red imported fire ant - Wikipedia
| Қызыл импортталған өрт құмырсқасы | |
|---|---|
 | |
| Өрт сөндіретін құмырсқалар тобы | |
| Ғылыми классификация | |
| Корольдігі: | Анималия |
| Филум: | Артропода |
| Сынып: | Инсекта |
| Тапсырыс: | Гименоптера |
| Отбасы: | Формицидалар |
| Субфамилия: | Мирмициналар |
| Тұқым: | Соленопсис |
| Түрлер: | S. invicta |
| Биномдық атау | |
| Solenopsis invicta Бүрэн, 1972 | |
| Синонимдер[1] | |
| |
The импортталған қызыл құмырсқа (Solenopsis invicta) деп те аталады от құмырсқасы немесе РИФА, Оңтүстік Америкадан шыққан құмырсқалардың бір түрі. Тұқым мүшесі Соленопсис подфамилияда Мирмициналар, ол болды сипатталған Швейцариялық энтомолог Феликс Санцчи 1916 жылғы нұсқа ретінде. Оның ағымы нақты аты инвикта құмырсқаға 1972 жылы жеке түр ретінде берілген. Алайда, нұсқасы мен түрлері бірдей құмырсқа болды және оның кең қолданылуының арқасында атау сақталды. Қызыл Америкадан шыққан қызыл отты құмырсқа Австралияда, Жаңа Зеландияда, бірнеше Азия мен Кариб теңізі елдерінде және Америка Құрама Штаттарында кездейсоқ енгізілген. Қызыл импортталған өрт құмырсқасы полиморфты, өйткені жұмысшылар әртүрлі формада және мөлшерде пайда болады. Құмырсқаның түсі қызыл, қоңыр немесе қара түске боялған газ, бірақ ерлер толығымен қара. Қызыл түсті импортталған өрт құмырсқалары өзгертілген аймақтарда басым және әр түрлі тіршілік ету ортасында тұрады. Олар жаңбырлы ормандарда, бұзылған жерлерде, шөлдерде, шабындықтарда, жолдар мен ғимараттардың жанында және электр жабдықтарында кездеседі. Колониялар топырақтан салынған үлкен қорғандар құрайды, себебі олар кіреберісі көрінбейді, өйткені жемдік тоннельдер салынып, жұмысшылар ұядан алыста пайда болады.
Бұл құмырсқалар әр түрлі мінез-құлықты көрсетеді, мысалы, су деңгейі көтеріліп жатқанын сезгенде сал салу. Олар сондай-ақ көрсетеді некрофориялық мінез-құлық, ұяшықтар ұядан тыс қоқыс үйінділеріндегі сынықтарды немесе өлі құмырсқаларды тастайды. Жемдеу жылы немесе ыстық күндерде жүреді, бірақ олар түнде сыртта қалуы мүмкін. Жұмысшылар бірқатар арқылы байланысады жартылай химия және феромондар жалдау, қоректену және қорғаныс үшін қолданылады. Олар жейтіндер және өлі сүтқоректілерді, буынаяқтыларды, жәндіктерді, тұқымдарды және тәтті заттарды жеу шіре бал бастап гемиптеран олар дамыған жәндіктер қатынастар. Жыртқыш аңдарға арахнидтер, құстар және көптеген жәндіктер, соның ішінде басқа құмырсқалар, инеліктер, құлаққаптар және қоңыздар жатады. Құмырсқа - паразиттер мен бірқатар патогенді, нематодалар мен вирустардың иесі, оларды ықтимал биологиялық бақылау агенттері ретінде қарастырды. Үйлену рейсі жылы мезгілдерде болады, ал алатес 30 минут бойы жұптасуы мүмкін. Колонияны құруды жалғыз патшайым немесе патшайымдар тобы жүзеге асыра алады, кейінірек алғашқы жұмысшылар пайда болғаннан кейін үстемдікке таласады. Жұмысшылар бірнеше ай, ал патшайымдар бірнеше жыл өмір сүре алады; колония саны 100000-нан 250000-ға дейін өзгеруі мүмкін. Қызыл импортталған өрт құмырсқасында қоғамның екі түрі бар: полигинді колониялар (көп ханшалы ұялар) және моногинозды колониялар (бір патшайым бар ұялар).
Құмырсқаның тіршілігінде улар маңызды рөл атқарады, өйткені ол олжа алу үшін немесе қорғаныс үшін қолданылады.[2] Удың 95% -ы суда ерімейтін заттардан тұрады пиперидин алкалоидтар ретінде белгілі соленопсиндер, қалғандары сезімтал адамдарға ерекше әсер етуі мүмкін улы белоктардың қоспасынан тұрады. Құрама Штаттарда олардан жыл сайын 14 миллионнан астам адам зардап шегеді, мұнда көптеген адамдар уға аллергия тудырады деп күтілуде. Зардап шеккендердің көпшілігінде қатты жану мен ісіну пайда болады, содан кейін стерильді пайда болады пустулалар, бұл бірнеше күн бойы қалуы мүмкін. Алайда адамдардың 0,6% -дан 6,0% -на дейін зардап шегуі мүмкін анафилаксия, бұл емделмеген жағдайда өлімге әкелуі мүмкін. Жалпы симптомдарға бас айналу, кеудедегі ауырсыну, жүрек айну, қатты тершеңдік, қан қысымының төмендеуі, ентігу және сөйлеудің бұзылуы жатады. Қызыл импортталған өрт құмырсқаларының шабуылынан 80-ден астам өлім тіркелді. Емдеу симптомдарға байланысты; тек ауырсыну мен пустула пайда болатындарға медициналық көмек қажет емес, бірақ анафилаксиядан зардап шегетіндерге көмек көрсетіледі эпинефриндер. Дененің тұтас сығындысы иммунотерапия құрбандарды емдеу үшін қолданылады және тиімділігі жоғары болып саналады.[дәйексөз қажет ]
Құмырсқа зиянды жәндік ретінде қарастырылады, ол жыл сайын миллиардтаған доллар шығын келтіріп, жабайы табиғатқа әсер етеді. Құмырсқалар қалалық жерлерде жақсы өседі, сондықтан олардың болуы ашық ауада өткізуге кедергі келтіруі мүмкін. Ұялар тротуарлар мен іргетастар сияқты құрылымдардың астына салынуы мүмкін, бұл құрылымдық проблемаларды тудыруы немесе құлап қалуы мүмкін. Олар құрылымдарды зақымдауы немесе бұзуы ғана емес, сонымен қатар қызыл импортталған өрт құмырсқалары жабдық пен инфрақұрылымды зақымдауы және бизнеске, жер мен мүлік құндылықтарына әсер етуі мүмкін. Ауыл шаруашылығында олар егін мен техниканы зақымдауы және жайылымға қауіп төндіруі мүмкін. Олар әр түрлі дақылдарды басып алатыны белгілі, ал ауылшаруашылық жерлеріне салынған қорғандар егін жинауға кедергі келтіруі мүмкін. Сондай-ақ олар жануарлар мен малдарға қауіп төндіреді, олар ауыр жарақат келтіруі немесе оларды өлтіруі мүмкін, әсіресе жас, әлсіз немесе ауру жануарларды. Осыған қарамастан, олар пайдалы болуы мүмкін, өйткені олар егінге қарапайым зиянкестер жәндіктерін пайдаланады. Осы құмырсқалармен күресудің кең таралған әдістеріне жем және фумигация жатады; басқа әдістер тиімсіз немесе қауіпті болуы мүмкін. Белгілі және маңызды болғандықтан, құмырсқа планетада ең зерттелген жәндіктердің біріне айналды, тіпті онымен қарсылас болды батыс бал арасы (Apis mellifera).[3][4]
Этимология және жалпы атаулар
The нақты эпитет қызыл импортталған өрт құмырсқасының, инвикта, туындайды Латын, және «жеңілмейтін» немесе «жеңілмеген» дегенді білдіреді.[5][6][7] Эпитет сөз тіркесінен бастау алады Рома инвикта дейін «шабыттандырылмаған Рим», шабыттандырушы дәйексөз ретінде қолданылған Батыс Рим империясының құлауы 476 жылы AD. The жалпы аты, Соленопсис, «сыртқы көрініс» немесе «бет» деп аударылады Ежелгі грек. Бұл қосылыс екі ежелгі грек сөзінен тұрады -солен, «құбыр» немесе «арна», және опсис, «көріну» немесе «көру» деген мағынаны білдіреді.[8][9] Құмырсқа «қызыл импортталған өрт құмырсқасы» деп аталады (қысқарған RIFA), себебі оның жануы сезімнің әсерінен пайда болады.[10][11] Балама атауларға мыналар жатады: «от құмырсқасы», «қызыл құмырсқа» немесе «қаңғыбас құмырсқа».[12][13] Бразилияда жергілікті тұрғындар құмырсқаны атайды toicinhera, бұл португал сөзінен шыққан toicinho (шошқа майы).[14]
Таксономия

Қызыл түсті импортталған өрт құмырсқасын алғаш рет швейцариялық энтомолог сипаттаған Феликс Санцчи жариялаған 1916 жылғы журнал мақаласында Физ.[15] Алғашында аталған Solenopsis saevissima wagneri а синтип бастап жиналған жұмысшы Сантьяго-дель-Эстеро, Аргентина, Санцки құмырсқаның нұсқасы деп санады S. saevissima; нақты эпитет, вагери, алғашқы үлгілерді жинаған Э.Р.Вагнердің тегінен шыққан.[16] The түрі материал қазіргі уақытта орналастырылған Naturhistorisches мұражайы Базель, Швейцария, бірақ қосымша типтегі жұмысшылар үйге орналастырылуы мүмкін Naturelle ұлттық музыкалық музейі, Париж.[17] 1930 жылы американдық мирмеколог Уильям Крейтон түріне шолу жасады Соленопсис және таксонды қайта жіктеді Solenopsis saevissima electra wagneri кезінде түршелік ол Санцкидің алғашқы сипаттамасына сілтеме жасаған бірде-бір жұмысшыны жинай алмайтынын атап өтті.[18] 1952 жылы S. saevissima түрлер кешені зерттелді және тоғыз басқа түр атауларымен бірге, S. saevissima electra wagneri синониміне ие болды S. saevissima saevissima.[19] Бұл қайта жіктеуді австралиялық энтомолог Джордж Эттершанк өзінің түрін қайта қарау кезінде және Вальтер Кемпфтің 1972 жылғы неотропикалық құмырсқалар каталогында қабылдады.[20][21]
1972 жылы американдық энтомолог Уильям Бурен жаңа түр деп санап, оны атады Solenopsis invicta.[22] Бүрен а голотип жұмысшы Куяба жылы Mato Grosso, Бразилия және Джорджия энтомологиялық қоғамы шығарған журнал мақаласында құмырсқаның алғашқы ресми сипаттамасын ұсынды. Ол кездейсоқ қате жіберіп алды инвикта сияқты инвика түрдің сипаттама беттерінің үстінде, дегенмен бұл анық болды инвикта мақалада атау үнемі қолданылғандықтан, емле мақсаты болған.[23] Материал қазіргі уақытта Ұлттық табиғи тарих мұражайы, Вашингтон, Колумбия округі[22]

1991 жылы түрлер кешеніне шолу жасаған кезде американдық энтомолог Джеймс Трэйджер синонимдес S. saevissima electra wagneri және S. wagneri бірге.[23] Трейгер қате сілтеме жасайды Solenopsis saevissima electra wagneri атау деп қате сеніп, түпнұсқа аты ретінде S. wagneri қол жетімсіз болды және Бүреннің атын қолданды S. invicta. Трейджер бұған дейін сенген S. invicta ерекше болды S. saevissima материалды салыстырғанға дейін S. wagneri. Трейджер бұл туралы айтады S. wagneri бар басымдық аяқталды S. invicta, бұл атау ешқашан түршелік дәрежеден жоғары қолданылған емес. Санцки кезінен бастап бұл атауды қолдану жинақталған үлгілермен байланысты емес, нәтижесінде номен нудум.[23] 1995 жылы ағылшын мирмеколог Барри Болтон танып, трагердің қатесін түзеді S. wagneri жарамды атау және синоним ретінде S. invicta.[24] Ол Трагерді қате жіктеген деп мәлімдейді S. wagneri қол жетімсіз атау және сілтемелер ретінде S. saevissima electra wagneri бастапқы таксон ретінде. Ол мұны аяқтайды S. wagneri шын мәнінде түпнұсқа атауы және артықшылығы бар S. invicta.[24][25]
1999 жылы Стив Шаттак және оның әріптестері бұл есімді сақтауды ұсынды S. invicta.[17] Бірінші сипаттамасынан бастап S. invictaБұл атауды қолданған 1800-ден астам ғылыми еңбектер оның экологиялық мінез-құлқы, генетикасы, химиялық байланысы, экономикалық әсерлері, бақылау әдістері, популяциясы және физиологиясы туралы көптеген тақырыптарды талқылады. Олар пайдалану деп мәлімдейді S. wagneri ғалымдар мен ғалым емес адамдарға қатысты номенклатуралық тұрақтылыққа «қауіп»; таксономистер мұндай атауды өзгертуге бейімделе алған болуы мүмкін, бірақ егер мұндай жағдай орын алса, есімдердің шатасуы пайда болуы мүмкін. Осыған байланысты Шаттук және оның әріптестері оны әрі қарай қолдануды ұсынды S. invicta және емес S. wagneri, өйткені бұл атау сирек қолданылған; 1995-1998 жылдар аралығында 100-ден астам мақала жарияланды S. invicta және тек үшеуі қолданады S. wagneri. Олар сұрады Зоологиялық номенклатура бойынша халықаралық комиссия (ICZN) пайдалану жалпы өкілеттіктер басу S. wagneri үшін емес, басымдық қағидаты үшін Омонимия принципі. Сонымен қатар, олар бұл атауды сұрады S. invicta зоологиядағы нақты атаулардың ресми тізіміне қосылады S. wagneri зоологиядағы қабылданбаған нақты атаулардың ресми индексіне қосылады.[17] Қарау кезінде ұсынысқа энтомологиялық қоғамдастық дауыс берді және оны бір сайлаушыдан басқалары қолдады. Олар басудың ешқандай негізі жоқ екенін атап өтті S. wagneri; оның орнына артықшылық берген дұрыс болар еді S. invicta аяқталды S. wagneri автор әрқашан оларды ерекше деп қарастырған кезде. ICZN сақтайды S. invicta және басу S. wagneri 2001 жылғы шолуда.[26] Осы классификацияға сәйкес, қызыл импортталған өрт құмырсқасы тұқымдас бөлігі болып табылады Соленопсис тайпада Solenopsidini, субфамилия Мирмициналар. Бұл отбасы мүшесі Формицидалар, бұйрыққа жататын Гименоптера, құрамында құмырсқалар бар жәндіктер қатары, аралар, және аралар.[27]


Филогения
Қызыл импортталған өрт құмырсқасы S. saevissima түр-топ. Мүшелерді жұмысшылар мен патшайымдардағы фуникулдың соңында екі біріктірілген клубтарымен ажыратуға болады, ал фундукуланың екінші және үшінші сегменттері үлкен жұмысшыларда екі есе ұзын және кең. Полиморфизм барлық түрлерінде кездеседі және төменгі жақ сүйектері төрт тісті көтеру.[22] Келесісі кладограмма қызыл импортталған өрт құмырсқасының басқа мүшелер арасындағы орнын көрсетеді S. saevissima түр-топ:[a][28]
| Соленопсис |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Фенотиптік және генетикалық деректер қызыл импортталған өрт құмырсқасы мен қара импортталған өрт құмырсқасы (Solenopsis richteri) бір-бірінен ерекшеленеді, бірақ олар жақын генетикалық байланыста болады.[29][30][31][32] Гибридизация екі құмырсқаның арасында олар байланысатын жерлерде, Миссисипиде орналасқан гибридті аймақпен кездеседі. Мұндай будандастыру осы екі құмырсқаның бірнеше онжылдықтар бұрын оңтүстікте алғаш кездескен кездегі екіншілік байланысынан туындады. Алабама.[29][33] Негізделген митохондриялық ДНҚ, қаралды гаплотиптер қалыптастырмаңыз монофилетикалық қаптау. Тексерілген гаплотиптердің кейбіреулері жақын қарым-қатынасты қалыптастырады S. мегергейттер, S. quinquecuspis және S. richteri басқаларымен салыстырғанда S. invicta гаплотиптер. Мүмкін болатын жағдай парафилетикалық топтау қызыл импортталған өрт құмырсқасы және S. quinquecuspis морфологиялық тұрғыдан ажырата алмайтын бірнеше түрден тұратын құпия түр топтары.[32][34]
Генетика
Зерттеулер көрсеткендей митохондриялық ДНҚ вариация айтарлықтай түрде пайда болады полигине қоғамдар (бірнеше патшайымдары бар ұялар),[35] бірақ ешқандай өзгеріс анықталған жоқ моногине қоғамдар (жалғыз ханшайымы бар ұялар).[36] Триплоидия (хромосомалық аномалия) қызыл импортталған өрт құмырсқаларында жоғары жылдамдықпен жүреді (репродуктивті емес аналықтарда 12% дейін), бұл диплоидты еркектердің жоғары жиілігімен байланысты.[37] Қызыл импортталған өрт құмырсқасы - бұл а-ға ие алғашқы түр жасыл сақалды ген, сол арқылы табиғи сұрыптау артықшылық бере алады альтруистік мінез-құлық. Бұл генді қамтитын жұмысшылар иесі бар аналықтарды, ал иелері жоқ белгілерді иіс белгілерін қолдану арқылы ажырата алады. Жұмысшылар гені жоқ патшайымдарды өлтіреді.[38][39] 2011 жылы ғалымдар бұл туралы жариялады толық тізбектелген қызыл импортталған өрт құмырсқасы геном еркектен.[40]
Сипаттама

Қызыл түсті импортталған өрт сөндірушілері олардың мөлшерін кішіден ортаға дейін өзгертеді полиморфты. Жұмысшылардың өлшемдері 2,4-тен 6,0 мм-ге дейін (0,094 және 0,236 дюйм).[41] Бастың өлшемдері 0,66 - 1,41 мм (0,026 - 0,056 дюйм), ені 0,65 - 1,43 мм (0,026 - 0,056 дюйм). Ірі жұмысшыларда (негізгі жұмысшылардағыдай) олардың бастарының ені 1,35 - 1,40 мм (0,053 - 0,055 дюйм) және 1,39 - 1,42 мм (0,055 - 0,056 дюйм). Антенна суреттер 0,96 - 1,02 мм (0,038 - 0,040 дюйм) және кеуде ұзындығы 1,70 - 1,73 мм (0,067 - 0,068 дюйм).[22] Көздің артында басы дөңгеленіп кеңейе түседі желке бөлімдері қазіргі және ұқсастықтан айырмашылығы S. richteri, лобтар ортаңғы сызықтан гөрі шыңға шығады, бірақ желке тәрізді эксцизия бүгілуге ұқсас емес. Ірі жұмысшылардың сызбалары желке шыңынан бір-екі диаметрге аспайды; бұл функция көбірек байқалады S. richteri. Орташа өлшемді жұмысшыларда сүйек шыңына жетеді және ең кішкентай жұмысшыларда артқы шекарадан асып түседі. Шағын және орта жұмысшыларда бас эллипс жағына ие болады. Кішкентай жұмысшылардың басы артқа қарағанда кеңірек.[22] Негізгі жұмысшыларда pronotum бұрыштық иықтары да, батып кеткен постеромедиан аймағы да жоқ. Промесонотум дөңес және продеум негізі дөңгелектелген, сонымен қатар дөңес. Негізі мен икемділігі бірдей ұзындықта болады. The тігіс Promesonotum үлкен жұмысшыларда күшті немесе әлсіз. The жапырақ қалың және доғал шкаласы бар; егер артынан байқалса, ол жоғарыда көрсетілгеннен гөрі дөңгелектелген емес S. richteri, ал кейде ол субтрунт болуы мүмкін. Постпетиола үлкен және кең, ал үлкен жұмысшыларда оның ұзындығынан кеңірек болады. Постпетиол алдыңғы жағынан азырақ және артқы жағынан кеңірек болады. Арқа бетінің артқы жағында көлденең әсер пайда болады. Жылы S. richteri, бұл функция да бар, бірақ әлдеқайда әлсіз.[22]
Мүсін өте ұқсас S. richteri.[22] Пункциялар пилотия пайда болатын жерден болады, және олар көбінесе бастың доральді және вентральды бөліктерінде созылады. Кеуде қуысында стриялар бар, бірақ олар азырақ ойылған, тесілгенге қарағанда азырақ S. richteri. Жапырақта нүктелер бүйірінде орналасқан. Постпетиола жоғарыда қарастырылған кезде айқын көлденең пунктостриялары бар күшті шағранға ие. Бүйір терең тесіктермен жабылған, олар кішірек, бірақ тереңірек көрінеді. Жылы S. richteri, тесіктері үлкен және таяз. Бұл бетіне мөлдір емес көрініс береді. Кейбір жағдайларда артқы бөліктің айналасында пунктуация болуы мүмкін.[22] Пилотность ұқсас болып көрінеді S. richteri. Бұл түктер тік және ұзындықтары әр түрлі, әр жағынан ұзын болып көрінеді pronotum және мезонотум; басында ұзын түктер бойлық қатарда көрінеді. Көптеген басылған түкті петиолар шкаласында; бұл керісінше S. richteri, өйткені бұл шаштар сирек.[22] Жұмысшылар қызыл немесе сәл сарғыш болып қоңыр немесе толық қара болып көрінеді газ.[15] Асқазан дақтары кейде үлкен жұмысшыларда байқалады, олар ондағыдай ашық түсті емес S. richteri. Асқазан дақтары әдетте бірінші асқазанның аз бөлігін жабады тергит. Кеуде қуысы консолорлы, ашық қызыл-қоңырдан қара-қоңырға дейін. Аяқтар және коксалар әдетте жеңіл көлеңкеленген. Үлкен жұмысшыларда бастың түсі сәйкес келеді желке және шың қоңыр болып көрінеді. Бастың басқа бөліктері, соның ішінде алдыңғы, гена және орталық аймақ клипей, не сарғыш, не сары-қоңыр. Геналар мен төменгі жақ сүйектерінің алдыңғы шекаралары қара-қоңыр түсті; сонымен қатар олардың екеуі де желкелермен бірдей түсті көлеңкеге ие көрінеді. Кескіндер және funiculi түсі баспен бірдей болады немесе желкемен желке бөледі. Кішкентай және орташа жұмысшылардағы бастың ашық түсті аймақтары тек маңдай аймағында шектеледі, мұнда жебеге немесе ракетаға ұқсас қараңғы белгі бар. Кейде ұялардың әр түрлі түстері болуы мүмкін. Мысалы, жұмысшылар әлдеқайда қараңғы, ал асқазан дақтары мүлдем болмауы немесе қара-қоңыр болып көрінуі мүмкін.[22]
Патшайымдардың бас ұзындығы 1,27 - 1,29 мм (0,050 - 0,051 дюйм), ені - 1,32 - 1,33 мм (0,052 - 0,052 дюйм).[22] Көлемдер 0,95 - 0,98 мм (0,037 - 0,039 дюйм), ал кеуде қуысы 2,60 - 2,63 мм (0,102 - 0,104 дюйм) құрайды. Басы дерлік ажыратылмайды S. richteri, бірақ оксипитальды экскизия қыртыстарға ұқсас емес, ал қабыршақтар айтарлықтай қысқа. Оның петиолды шкаласы дөңес және масштабқа ұқсас S. richteri. Постпетиоланың түзу жақтары бар, олар ешқашан ойыспайды S. richteri олар ойысатын жерде. Кеуде қуысы іс жүзінде бірдей, бірақ метаплевралық стриат аймағы мен продееал арасындағы бос кеңістік спирактар тар мыжылған немесе жоқ. Жапырақтың бүйір бөліктері нүктелі. Постпетиолдың бүйір жақтары мөлдір емес тесіктері бар, бірақ дұрыс емес кедір-бұдыр көрінбейді. Дорсумның алдыңғы бөлігі шағыл және ортаңғы және артқы аймақтар көлденең пункто-стрияларға ие. Осы аймақтардың барлығында тік шаштар бар. Петиолдің де, постпетиолдың да алдыңғы бөліктері продедеумда байқалатын жасушалық өсіндіге ие. Патшайымның түсі жұмысшының түсіне ұқсас: газатор қара қоңыр, ал аяқтары, қабыршақтары және кеудесі ашық қоңыр, қара жолақтары бар мезоскутум. Орталық аймақтардың айналасында басы сарғыш немесе сарғыш-қоңыр, желке және төменгі жақ сүйектері кеуде қуысына ұқсас, ал қанат тамырлары түссізден бозғылт қоңырға дейін созылады.[22] Еркектері ұқсас S. richteri, бірақ петиолды қабыршақтың жоғарғы шекаралары ойыс болады. Екі түрде де постпетиол мен петиол спиралдары қатты проекциялайды. Ер адамның бүкіл денесі түсті қара түсті, бірақ антенналары ақшыл. Патшайым сияқты қанат тамырлары түссіз немесе ақшыл қоңыр болады.[22]

Қызыл импортталған құмырсқаны ұқсас түрдегі қате деп анықтауға болады S. richteri.[22] Екі түрді бір-бірінен бас, кеуде қуысы және постпетиолды морфологиялық зерттеу арқылы ажыратуға болады. Жылы S. richteri, бастың бүйірлері кең эллипс тәрізді және қызыл түсті импортталған өрт құмырсқасында байқалатын кордалық пішін жоқ. Ортаңғы сызық пен желке экскизиясының жанында орналасқан желке лобтарының аймағы мыжылған тәрізді көрінеді S. richteri қызыл импортталған отты құмырсқаға қарағанда. туралы Марина S. richteri қызыл импортталған өрт құмырсқасына қарағанда ұзын, ал пронотумның бұрыштық иықтары күшті. Мұндай сипат қызыл импортталған өрт құмырсқасында жоқтың қасы. Таяз, бірақ батып кеткен аймақ тек үлкен жұмысшыларда белгілі S. richteri, артқы аймағында орналасқан дорсум Пронотум. Бұл функция қызыл импортталған қызыл өрт сөндірушілерінде мүлдем жоқ. Қызыл импортталған өрт құмырсқасының промесонотасы қатты дөңес, ал бұл қасиеті әлсіз дөңес S. richteri. Қарап тексергенде пропедаумның негізі созылып, түзу орналасқан S. richteriқызыл, ал дөңес және қысқа импортталған өрт құмырсқасы. Сондай-ақ, оның тік немесе әр түрлі жақтары бар кең постпетиол бар. Постпетиол S. richteri жақтары жақындатылған тар. Жылы S. richteri, постпетиолдың постеродоральды бөлігіндегі көлденең әсер күшті, бірақ қызыл импортталған өрт құмырсқасында әлсіз немесе жоқ.[22] Сонымен қатар, S. richteri жұмысшылар қызыл импортталған өрт құмырсқалары жұмысшыларынан 15% үлкен, қара-қоңыр және газердің артқы жағында сары жолақ бар.[42][43]
Брод

Жұмыртқалар кішкентай және сопақ тәрізді, олардың мөлшері шамамен бір аптаға дейін сақталады. Бір аптадан кейін жұмыртқа эмбрионның формасын қабылдайды және жұмыртқа қабығын алып тастаған кезде личинка түрінде қалыптасады.[44] Личинкалардың өлшемі 3 мм (0,12 дюйм).[45] Олар ұқсас көріністі көрсетеді S. geminata дернәсілдер, бірақ оларды интеграл арқылы ажыратуға болады жұлын артқы сомиттердің доральды бөлігінің жоғарғы жағында. Дене түктері 0,063-тен 0,113 мм-ге дейін (0,0025-тен 0,0044 дюймге дейін) дентикулат ұшымен өлшенеді. Антенналардың екеуінде де екі-үшеу бар сенсилла. The лабрум алдыңғы бетінде 0,013 мм (0,00051 дюйм) екі түктерімен кішірек. The жоғарғы жақ сүйегі карточка мен стипендия арасында склеротирленген жолақ бар. The лабиум сонымен қатар шағын склеротизацияланған жолағы бар.[45] Түтіктері ерін бездері ас қорыту ферменттерінің мол деңгейіне ие ақуызды зат шығаратыны немесе шығаратыны белгілі, оған қатты тағамның ішектен тыс қорытылуы ретінде қызмет ететін протеазалар мен амилазалар кіреді. Ортаңғы ішекте де бар амилазалар, roteases және upases. Оның су қоймасындағы тар клеткалардың секреция кезінде атқаратын қызметтері шамалы.[46] Қуыршақтар кез-келген кастаның ересектеріне ұқсайды, тек олардың аяқтары мен антенналары денеге мықтап ұсталады. Олар ақ болып көрінеді, бірақ уақыт өте келе, қуыршақтар пісуге дайын болғанда қараңғы болады.[47]
Төрт личинка instars ерекше морфологиялық белгілерге сүйене отырып сипатталған.[44][48][49] Кәмелетке толмаған және негізгі жұмыскерлердің личинкаларын соңғы айырмашылыққа дейін, өлшем айырмашылықтары айқын болған кезде ажырату мүмкін емес.[49] Қуыршақ кезінде касталар арасындағы бас енінің айырмашылығы кеңірек болады. Репродуктивті дернәсілдер жұмысшы дернәсілдеріне қарағанда үлкенірек және ауыз тістеріндегі дискретті морфологиялық айырмашылықтарды көрсетеді.[49] Төртінші ерлер мен аналықтардың личинкаларын олардың пішіні мен дене түсіне байланысты ажыратуға болады,[49] және ішкі гоноподальды қиял дискілері ерекшеленуі мүмкін.
Полиморфизм
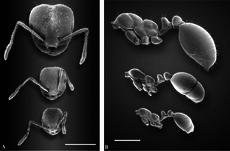
Қызыл түсті импортталған өрт құмырсқасы екі түрлі жұмысшылар кастасымен полиморфты: кішігірім жұмысшылар және ірі жұмысшылар (сарбаздар). Полиморфизмді көрсететін көптеген құмырсқалар сияқты, жас, кішігірім құмырсқалар жем болмайды және керісінше, үлкен жұмысшылар сыртқа шығып, жемшөп алады.[50][51][52][53] Бастапқы колонияда полиморфизм жоқ, бірақ оның орнына оларды «минимум» немесе «нанитика» деп аталатын мономорфты жұмысшылар алады. Сыналған колониялардағы бастың орташа ені дамудың алғашқы алты айында өседі.[54][55] Бес жасар колонияларда кәмелетке толмаған жұмысшылардың бас ені азаяды, бірақ негізгі жұмысшылар үшін бас ені өзгеріссіз қалады. Негізгі жұмысшының жалпы салмағы кәмелетке толмаған жұмысшының олар келген кездегіден екі есе артық, ал алты айға қарай негізгі жұмысшылар кәмелетке толмаған жұмысшыларға қарағанда төрт есе ауыр. Ірі жұмысшылар дамығаннан кейін, олар жұмыс күшінің көп бөлігін құра алады, олардың 35% -ы бір колониядағы негізгі жұмысшылар.[54] Бұл колонияның өнімділігіне әсер етпейді, өйткені кішкентай жұмысшылармен полиморфты колониялар мен ұялар шамамен бірдей жылдамдықта балаларды шығарады, ал тамақ көздері шектелмеген кезде полиморфизм артықшылығы немесе кемшілігі болмайды. Алайда, полиморфты колониялар энергиялық жағынан тиімдірек, ал азық-түлік шектеулі болған жағдайда, полиморфизм балапандар өндірісінде кішкене артықшылықтар беруі мүмкін, бірақ бұл тағамдық стресстің деңгейіне байланысты.[56]
Жұмысшы құмырсқалар үлкен мөлшерге дейін өскен сайын бастың ұзындығы дененің жалпы ұзындығымен бір уақытта өсуіне байланысты бастың формасы өзгереді, ал бас ені 20% өсуі мүмкін. Антенналардың ұзындығы тек баяу өседі; дененің ұзындығы екі есе артқан кезде антенналар тек 60% ұзаруы мүмкін, сондықтан дененің ұзындығы екі еселенген сайын антеннаның салыстырмалы ұзындығы 20% -ға азаяды.[55] Дененің барлық жеке аяқтары изометриялық болып табылады, яғни дененің ұзындығы екі еселенген кезде де аяқтар екі еселенеді. Алайда, аяқтардың барлығы бірдей ұзын емес; проторакалық бөлік аяғының ұзындығының 29%, мезоторакалы 31%, ал метаторастық 41% құрайды. Алғашқы екі жұп аяқтың ұзындығы бір-біріне тең, ал соңғы жұп ұзынырақ.[55] Тұтастай алғанда, жұмысшының морфологиялық келбеті ол үлкейгенде күрт өзгереді. Бас пішіннің ең үлкен өзгеруін және биіктігін көрсетеді алинотум ұзындығынан тез өседі, мұнда кәмелетке толмаған жұмысшыларда 0,27 және негізгі жұмысшыларда 0,32 биіктік / ұзындық қатынасы көрінеді.[55] Осыған байланысты, үлкен жұмысшылар кішігірім жұмысшылардан айырмашылығы, кескінді және берік алинотумға ие. Дененің өлшемі өзгерген кезде ешқандай петит сегменті пішіннің өзгеруін көрсетпейді. Бензиннің ені оның ұзындығынан тез өседі, мұнда ені оның ұзындығының 96% құрауы мүмкін, бірақ 106% дейін өседі.[55]
Физиология
Басқа жәндіктер сияқты қызыл импортталған өрт құмырсқасы газ толтырылған түтіктер жүйесі арқылы тыныс алады трахеялар спирактар арқылы сыртқы ортаға байланысты. Трахея терминалдары (трахеолдар ) ішкі органдармен және тінмен тікелей байланысқа түсу. Оттегінің жасушаларға тасымалдануы (және жасушалардан шыққан көмірқышқыл газы) арқылы жүреді диффузия трахеолдар мен оны қоршаған тіндердің арасындағы газдардың және а үзіліссіз газ алмасу.[57] Басқа жәндіктер сияқты трахея жүйесі мен тіндердің арасындағы тікелей байланыс O-ны тасымалдау үшін сұйықтықтың айналымды желісінің қажеттілігін жояды.2.[58] Осылайша, қызыл импортталған өрт құмырсқалары мен басқа да буынаяқтылар қан айналымы жүйесінің қарапайым болуы мүмкін, дегенмен олар метаболизмге өте қымбат.[59]
The экскреторлық жүйе үш облыстан тұрады. Базальды аймақта ортаңғы ішектің артқы бөлігінде орналасқан үш жасуша бар. Алдыңғы және жоғарғы қуыстар төртеудің негіздерімен қалыптасады Мальпигия түтікшелері.[60] Жоғарғы қуыс люмен ащы ішектің. Тік ішек - бұл личинкалардың артқы бесінші бөлігін алып жатқан үлкен, бірақ жұқа қабырғалы қапшық. Қалдықтардың шығарылуы бақыланады ректальды клапандар бұл анусқа әкеледі.[60] Кейде, личинкалар құрамына сұйықтық бөледі зәр қышқылы, су және тұздар.[61] Бұл құрамды көбінесе жұмысшылар сыртқа шығарады және сыртқа шығарады, бірақ су стрессіндегі колониялар құрамын тұтынуы мүмкін.[60] Ішінде репродуктивті жүйе, ханшайымдар а феромон бұл мәмілеге кедергі келтіреді және оогенез қыздарда; колонияларда патшайымсыз тексерілгендер басталады ооцит мәміле жасасқаннан кейін дамыту және жұмыртқа салушы рөлге ие болу.[62] Ұшу бұлшықеттерінің деградациясы жұптасу және жасөспірім гормондары арқылы басталады және алдын алады корпус аллатэктомиясы.[63][64] Гистолиз ерігеннен басталады миофибрил және баяу бұзылуы миофиламенттер. Мұндай еру Z-сызығының жалғыз бос материалдарына жеткенге дейін жалғасады, олар да жоғалады; тек ядролар және пластиналы денелер қалу.[60] Бір зерттеуде аминқышқылдары ұлғаюы гемолимф ұрықтандырудан кейін.[65]
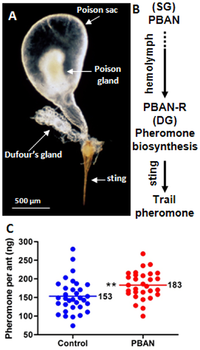
The бездер жүйесі құрамында төрт без бар: төменгі жақ, жақ, ерін және жұтқыншақтан кейінгі бездер.[60] Жұтқыншақтан кейінгі патшайым жақсы дамыған, ал басқа бездер жұмысшыларда үлкенірек. Жұтқыншақтан кейінгі без сіңіру үшін вакуум қызметін атқарады май қышқылдары және триглицеридтер, сонымен қатар асқазан ішек.[66] Басқа бездердің қызметі нашар зерттелген болып қалады. Ересек құмырсқалардың ас қорыту жүйесінің ферменттерін талқылаған бір зерттеуде, липаза белсенділігі төменгі жақ және ерін бездерінде, сонымен қатар анықталды инвертаза белсенділік. The Дюфур безі табылған құмырсқа көзі болып табылады феромондар Дегенмен, ғалымдар улы безі феромон патшайымының көзі деп санады.[60][67][68] Нейрогормон нейропептидті белсендіретін феромон биосинтезі Дюфур безінен феромондардың биосинтезін белсендіретін құмырсқада кездеседі.[69] The сперматека без сперматозоидтар қызметін атқаратын аналықтарда кездеседі. Еркектерде бұл бездер жетіспейтін сияқты, бірақ оның басымен байланысты адамдар морфологиялық тұрғыдан жұмысшыларда кездеседі, бірақ бұл бездер басқаша әрекет етуі мүмкін.[66]

Құмырсқа қоршаған ортаның тыныс алуына байланысты көптеген тыныс алу проблемаларына тап болады, бұл олардың көбеюіне әкелуі мүмкін құрғау, гипоксия, және гиперкапния. Ыстық, ылғалды климат жүректің соғу жиілігін және тыныс алуды күшейтеді, демек, энергия мен судың шығыны артады.[58][70] Гипоксия мен гиперкапния желдетілмеген жерде тұратын қызыл импортталған отты құмырсқалар колониясынан туындауы мүмкін терморегуляция үйінділер мен жер астындағы ұялар. Үзіліссіз газ алмасу (DGE) құмырсқаларға өз шұңқырларында жиі кездесетін гиперкапникалық және гипоксиялық жағдайлардан аман қалуға мүмкіндік беруі мүмкін;[57] ол осы жағдайларға бейімделу үшін өте қолайлы, өйткені ол құмырсқалар О периодын ұлғайтуға мүмкіндік береді2 қабылдау және CO2 спираль манипуляциясы арқылы дербес шығару. Қызыл импортталған өрт құмырсқасының шабуылдағы жетістігі оның физиологиялық төзімділігімен байланысты болуы мүмкін абиотикалық стресс қарағанда ыстыққа төзімді және десекция стрессіне бейімделгіш S. richteri. Демек, құмырсқа жылу мен құрғау стрессіне аз ұшырайды. Дегенмен S. richteri қызыл импортталған өрт құмырсқасына қарағанда су құрамы жоғары, S. richteri кептіру стрессіне осал болды. Құрғауға сезімталдықтың төмендеуі судың төмендеуіне байланысты.[71] Көлеңкеленбеген және жылы жерлерде тұратын колонияларда көлеңкелі және салқын жерлерде тұратындарға қарағанда жылу төзімділігі жоғары болады.[72]
Тыныс алуға жанама әсер ететін зат алмасу жылдамдығына қоршаған ортаның температурасы да әсер етеді. Шың метаболизм шамамен 32 ° C температурада болады.[73] Метаболизм, демек, тыныс алу жылдамдығы температура жоғарылаған сайын тұрақты түрде жоғарылайды. DGE 25 ° C-тан жоғары деңгейде тоқтайды, бірақ оның себебі қазіргі кезде белгісіз.[74]
Тыныс алу жылдамдығына касталар да айтарлықтай әсер етеді. Еркектер әйелдер мен жұмысшыларға қарағанда тыныс алудың едәуір жоғары жылдамдығын көрсетеді, бұл ішінара олардың ұшуға қабілеттілігіне және бұлшықет массасының жоғарылауына байланысты. Жалпы алғанда, еркектерде бұлшықеттер көп және майлар аз, нәтижесінде метаболизм O жоғарылайды2 сұраныс.[74]Метаболизм жылдамдығы 32 ° C-та жоғары болса, колониялар көбінесе салқын температурада (25 ° C шамасында) жақсы дамиды. Температураның жоғарылауымен байланысты метаболикалық белсенділіктің жоғары жылдамдығы колония өсуінің шектеу факторы болып табылады, өйткені тамақ тұтыну қажеттілігі де жоғарылайды. Нәтижесінде үлкен колониялар салқын жағдайда болады, өйткені колонияны ұстап тұруға қажетті метаболизмге деген қажеттілік азаяды.[73]
Таралу және тіршілік ету аймағы
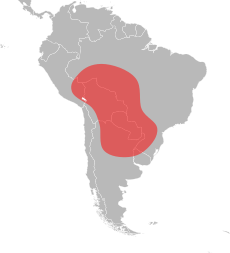
Қызыл түсті импортталған өрт құмырсқалары Орталық және Оңтүстік Американың тропикалық аймақтарына тән, олар Перудің оңтүстік-шығысынан Аргентинаның орталық бөлігіне және Бразилияның оңтүстігіне дейін созылатын кең географиялық диапазоны бар.[75][76][77][78] Оның Солтүстік Америкадағы географиялық диапазонынан айырмашылығы, Оңтүстік Америкадағы ауқымы айтарлықтай ерекшеленеді. Оның солтүстік-оңтүстік диапазоны өте ұзын, бірақ шығысы мен батысы өте тар таралған. Қызыл түсті импортталған қызыл құмырсқалардың солтүстік жазбалары Порту-Велхо Бразилияда, және оның оңтүстік рекорды болып табылады Қарсылық Аргентинада; бұл шамамен 3000 км (1900 миль) қашықтық. Салыстырмалы түрде, оның тар диапазонының ені шамамен 350 км (220 миль) құрайды және бұл Аргентина мен Парагвайдың оңтүстігінде және Амазонка өзенінің бассейнінің солтүстік аймақтарында тар болуы мүмкін.[79] Қызыл импортталған өрт құмырсқасының көптеген белгілі жазбалары айналасында Пантанал аймағы Бразилия. Алайда, бұл аймақтың іші мұқият зерттелмеген, бірақ түрдің оның айналасында қолайлы жерлерде кездесетіні анық. Пантанал аймағы қызыл импортталған от құмырсқасының түпнұсқа отаны болып саналады; гидрохордың таралуы қалқымалы құмырсқалар арқылы оңтүстік популяцияларды оңай есептей алатын Парагвай және Гуапере өзендері. Оның ауқымының батыс шегі нақты белгісіз, бірақ оның көптігі шектеулі болуы мүмкін. Пантанал аймағының болуына байланысты ол Боливияның шығысында кең болуы мүмкін.[79]
Бұл құмырсқалар Аргентинаның тумасы, ал қызыл импортталған отты құмырсқа, ең алдымен, олар Америка Құрама Штаттарына алғаш басып кірген кезде пайда болған; Атап айтқанда, осы құмырсқалардың популяциялары провинцияларынан табылды Чако, Корриентес, Формоза, Сантьяго-дель-Эстеро, Санта-Фе, және Тукуман.[23][79][80] Аргентинаның солтүстік-шығыс аймақтары - бұл құмырсқалардың басталатын ең сенімді болжамы.[77] Бразилияда олар Mato Grosso солтүстігінде және Рондония және Сан-Паулу штаты. Қызыл импортталған өрт құмырсқасы және S. saevissima болып табылады парапатриялық Бразилияда, байланыс аймақтары бар Mato Grosso do Sul, Парана штаты және Сан-Паулу.[76][81] In Paraguay they are found throughout the country, and have been recorded in Бокерон, Каагасу, Canindeyú, Орталық, Гуайра, Embeembucú, Парагуари, және Президент Хейз departments; Trager claims that the ant is distributed in all regions of the country.[82][83][84] They are also found in a large portion of northeastern Bolivia and, to a lesser extent, in northwestern Uruguay.[79][85]

The red imported fire ant is able to dominate altered areas and live in a variety of habitats. It can survive the extreme weather of the South American rain forest, and in disturbed areas, nests are seen frequently alongside roads and buildings.[86][87] The ant has been observed frequently around the жайылмалар of the Paraguay River.[88] In areas where water is present, they are commonly found around: irrigation channels, lakes, ponds, reservoirs, rivers, streams, riverbanks, and mangrove swamps.[82][86] Nests are found in agricultural areas, coastlands, wetlands, coastal dune remnants, deserts, forests, grasslands, natural forests, oak woodland, мезик орманы, leaf-litter, beach margins, shrublands, alongside rail and roads, and in urban areas.[89] In particular, they are found in cultivated land, managed forests and plantations, disturbed areas, intensive livestock production systems, and greenhouses.[86][90] Red imported fire ants have been found to invade buildings, including medical facilities.[91] In urban areas, colonies dwell in open areas, especially if the area is sunny.[91] This includes: urban gardens, picnic areas, lawns, playgrounds, schoolyards, parks, and golf courses.[82][91] In some areas, there are on average 200 mounds per acre.[92] During winter, colonies move under pavements or into buildings, and newly mated queens move into pastures.[86][91] Red imported fire ants are mostly found at altitudes between 5 and 145 m (16 and 476 ft) above sea level.[82]
Mounds range from small to large, measuring 10 to 60 cm (3.9 to 23.6 in) in height and 46 cm (18 in) in diameter with no visible entrances.[86][93] Workers are only able to access their nests thorugh a series of tunnels that protrude from the central region. Such protrusions can span up to 25 feet away from the central mound, either straight down in to the ground or, more commonly, sideways from the original mound.[94] Constructed from soil, mounds are oriented so that the long portions of the mound face toward the sun during the early morning and before sunset.[86][93] Mounds are usually oval-shaped with the long axis of the nest orientating itself in a north–south direction.[95] These ants also spend large amounts of energy in nest construction and transporting brood, which is related with терморегуляция. The brood is transported to areas where temperatures are high; workers track temperature patterns of the mound and do not rely on behavioural habits.[96] Inside nests, mounds contain a series of narrow horizontal tunnels, with subterranean shafts and nodes reaching grass roots 10 to 20 cm (3.9 to 7.9 in) below the surface; these shafts and nodes connect the mound tunnels to the subterranean chambers. These chambers are about 5 cm2 (0.77 inch2) and reach depths of 10 to 80 cm (3.9 to 31.5 in). The mean number of ants in a single subterranean chamber is around 200.[97][98][99]
Кіріспелер

Red imported fire ants are among the worst инвазиялық түрлер Әлемде.[100][101] Some scientists consider the red imported fire ant to be a "disturbance specialist"; human disturbance to the environment may be a major factor behind the ants' impact (fire ants tend to favour disturbed areas). This is shown through one experiment, demonstrating that mowing and plowing in studied areas diminished the diversity and abundance of native ant species, whereas red imported fire ants found on undisturbed forest plots had only diminished a couple of species.[102][103]
In the United States, the red imported fire ant first arrived in the seaport of Мобайл, Алабама, by cargo ship between 1933 and 1945.[b][c][79][106][107][108] Arriving with an estimated 9 to 20 unrelated queens,[109][110] the red imported fire ant was only rare at the time, as entomologists were unable to collect any specimens (with the earliest observations first made in 1942, preceded by a population expansion in 1937); the population of these ants exploded by the 1950s.[111][112][113] Since its introduction to the United States, the red imported fire ant has spread throughout the southern states and northeastern Mexico, negatively affecting wildlife and causing economic damage.[75][114][115][116] The expansion of red imported fire ants may be limited since they are almost wiped out during Теннесси winters, thus they may be reaching their northernmost range.[117][118][119] However, global warming may allow the red imported fire ant to expand its geographical range.[120] As of 2004, the ant is found in 13 states and occupies over 128 million hectares of land, and as many as 400 mounds can be found on a single acre of land.[121][122] The Америка Құрама Штаттарының Ауыл шаруашылығы министрлігі estimates that they expand 193 km (120 mi) per year.[91]
Red imported fire ants were first discovered in Квинсленд, Australia, in 2001.[123][124] The ants were believed to be present in shipping containers arriving at the Брисбен порты, most likely from North America.[125] Anecdotal evidence suggests fire ants may have been present in Australia for six to eight years prior to formal identification. The potential damage from the red imported fire ant prompted the Australian government to respond rapidly. A joint state and federal funding of $ A 175 million was granted for a six-year eradication programme.[126][127][128] Following years of eradication, eradication rates of greater than 99% from previously infested properties were reported. The program received extended Достастық funding of around A$10 million for at least another two years to treat the residual infestations found most recently.[129] In December 2014, a nest was identified at Порттық ботаника, Сидней, жылы Жаңа Оңтүстік Уэльс. The port was quarantined, and a removal operation took place.[130] In September 2015, populations originating from the United States were found at a Brisbane airport.[131]

Red imported fire ants have spread beyond North America. The Invasive Species Specialist Group (ISSG) reports the ant inhabiting from three of the Кайман аралдары. However, the sources the ISSG cited give no report about them on the island, but recent collections indicate that they are present.[75] In 2001, red imported fire ants were discovered in New Zealand, but they were successfully eradicated several years later.[75][132] Red imported fire ants have been reported in India,[133] Малайзия,[134] Филиппиндер[135] and Singapore.[75] However, these reports were found to be incorrect as the ants collected there were incorrectly identified as the red imported fire ant. In Singapore, the ants were most likely misidentified as well. In India, surveyed ants in Sattur Taluk, India listed the red imported fire ant there in high populations; meanwhile, no reports of the ant were made outside the surveyed area. In 2016, scientists state that despite no presence of the ant in India, the red imported fire ant will more than likely find suitable habitats within India's ecosystem if given the opportunity.[136] The reports in the Philippines most likely misidentified collected material as the red imported fire ant, as no populations have been found there.[75] Ол анықталды Гонконг and mainland China in 2004, where they have spread into several provinces as well as Макао және Тайвань.[137][138][139][140] No geographic or climatic barriers prevent these ants from spreading further, thus it may spread throughout the tropical and subtropical regions of Asia.[121][141] In Europe, a single nest was found in the Netherlands in 2002.[78]
Around 1980, red imported fire ants began spreading throughout the Батыс Үндістан, where they were first reported in Пуэрто-Рико және АҚШ-тың Виргин аралдары.[142][143] Between 1991 and 2001, the ant was recorded from Trinidad and Tobago, several areas in the Bahamas, the British Virgin Islands, Antigua, and the Turks and Caicos Islands.[75][144][145] Since then, red imported fire ants have been recorded on more islands and regions, with new populations discovered in: Ангилья, Әулие Мартин, Barbuda, Монтсеррат, Сент-Китс, Невис, Аруба, and Jamaica.[75] The ants recorded from Aruba and Jamaica have only been found on golf courses; these courses import сода бастап Флорида, so such importation may be an important way for the ant to spread throughout the West Indies.[75]
Populations found outside North America originate from the United States. In 2011, the DNA of specimens from Australia, China, and Taiwan was analysed with results showing that they are related to those in the United States.[146] Despite its spread, S. geminata has a greater geographical range than the red imported fire ant, but it can be easily displaced by it. Because of this, almost all of its exotic range in North America has been lost and the ant has almost disappeared there. On roadsides in Florida, 83% of these sites had S. geminata present when the red imported fire ant was absent, but only 7% when it is present.[75] This means that the ant can probably invade many tropical and subtropical regions where S. geminata populations are present.[75]
Мінез-құлық және экология

Red imported fire ants are extremely resilient and have adapted to contend with both flooding and drought conditions. If the ants sense increased water levels in their nests, they link together and form a ball or raft that floats, with the workers on the outside and the queen inside.[147][148][149] The brood is transported to the highest surface.[150] They are also used as the founding structure of the raft, except for the eggs and smaller larvae. Before submerging, the ants will tip themselves into the water and sever connections with the dry land. In some cases, workers may deliberately remove all males from the raft, resulting in the males drowning. The longevity of a raft can be as long as 12 days. Ants that are trapped underwater escape by lifting themselves to the surface using bubbles which are collected from submerged substrate.[150] Owing to their greater vulnerability to predators, red imported fire ants are significantly more aggressive when rafting. Workers tend to deliver higher doses of venom, which reduces the threat of other animals attacking. Due to this, and because a higher workforce of ants is available, rafts are potentially dangerous to those that encounter them.[151]
Necrophoric behaviour occurs in the red imported fire ant. Workers discard uneaten food and other such wastes away from the nest. The active component was not identified, but the fatty acids accumulating as a result of decomposition were implicated and bits of paper coated with synthetic oleic acid typically elicited a necrophoric response. The process behind this behaviour in imported red fire ants was confirmed by Blum (1970): unsaturated fats, such as oleic acid, elicit corpse-removal behaviour.[152] Workers also show differentiated responses towards dead workers and pupae. Dead workers are usually taken away from the nest, whereas the pupae may take a day for a necrophoric response to occur. Pupae infected by Metarhizium anisopliae are usually discarded by workers at a higher rate; 47.5% of unaffected corpses are discarded within a day, but for affected corpses this figure is 73.8%.[153]
Red imported fire ants have negative impacts on seed germination. The extent of the damage, however, depends on how long seeds are vulnerable for (dry and germinating) and by the abundance of the ants.[154] One study showed that while these ants are attracted to and remove seeds which have adapted for ant dispersal, red imported fire ants damage these seeds or move them in unfavourable locations for germination. In seeds given to colonies, 80% of Sanguinaria canadensi seeds were scarified and 86% of Viola rotundifolia seeds were destroyed.[155] Small percentages of longleaf pine (Pinus palustris) seeds deposited by workers successfully germinate, thus providing evidence that red imported fire ants help the movement of seeds in the longleaf pine ecosystem. Elaiosome-bearing seeds are collected at a higher rate in contrast to nonelaiosome-bearing seeds and do not store them in their nests, but rather in surface trash piles in the mound vicinity.[156]
Foraging and communication

Colonies of the red imported fire ant have tunneling surfaces that protrude out of the surfaces where workers forage.[157][158] These areas of protrusion tend to be within their own territory, but greater ant colonisation can affect this.[159] Tunnels are designed to allow effective body, limb and antennae interactions with walls, and a worker can also move exceptionally fast inside them (more than nine bodylengths per second).[160] The holes exit out of any point within the colony's territory, and foraging workers may need to travel half a metre to reach the surface. Assuming the average forager travels 5 m, over 90% of foraging time is inside the tunnels during the day and rarely at night. Workers forage in soil temperatures reaching 27 °C (80 °F) and surface temperatures of 12–51 °C (53–123 °F).[158] Workers exposed to temperatures of 42 °C (107 °F) are at risk of dying from the heat.[158] The rate of workers foraging drops rapidly by autumn, and they rarely emerge during winter. This may be due to the effects of soil temperature, and a decreased preference for food sources. These preferences only decrease when brood production is low. In the northern regions of the United States, areas are too cold for the ant to forage, but in other areas such as Florida and Техас, foraging may occur all year round. When it is raining, workers do not forage outside, as exit holes are temporarily blocked, pheromone trails are washed away, and foragers may be physically struck by the rain. The soil's moisture may also affect the foraging behaviour of workers.[158]
When workers are foraging, it is characterised by three steps: searching, recruitment, and transportation.[161] Workers tend to search for honey more often than other food sources, and the weight of food has no impact on searching time. Workers may recruit other nestmates if the food they have found is too heavy, taking as much as 30 minutes for the maximum number of recruited workers to arrive. Lighter food sources take less time and are usually transported rapidly.[161] Foraging workers become scouts and search solely for food outside the surface, and may subsequently die two weeks later from old age.[162]
Workers communicate by a series of semiochemicals және pheromones. These communication methods are used in a variety of activities, such as nestmate recruitment, foraging, attraction, and defence; for example, a worker may secrete trail pheromones if a food source it discovered is too large to carry.[163] These pheromones are synthesized by the Дюфур безі and may trail from the discovered food source back to the nest.[164][165] The components in these trail pheromones are also species-specific to this ant only, in contrast to other ants with common tail pheromones.[166] The poison sack in this species has been identified as being the novel storage site of the queen pheromone; this pheromone is known to elicit orientation in worker individuals, resulting in the deposition of brood.[68][167] It is also an attractant, where workers aggregate toward areas where the pheromone has been released.[163] A brood pheromone is possibly present, as workers are able to segregate brood by their age and caste, which is followed by licking, grooming and antennation.[163] If a colony is under attack, workers will release alarm pheromones.[168] However, these pheromones are poorly developed in workers. Workers can detect пиразиндер which are produced by the alates; these pyrazines may be involved in кешкі рейс, as well as an alarm response.[169]
Red imported fire ants can distinguish nestmates and non-nestmates through chemical communication and specific colony odours.[163][170] Workers prefer to dig into nest materials from their own colony and not from soil in unnested areas or from other red imported fire ant colonies. One study suggests that as a colony's diet is similar, the only difference between nested and unnested soil was the nesting of the ants themselves. Therefore, workers may transfer colony odour within the soil.[170] Colony odour can be affected by the environment, as workers in lab-reared colonies are less aggressive than those in the wild.[171] Queen-derived cues are able to regulate nestmate recognition in workers and амин деңгейлер. However, these cues do not play a major role in colony-level recognition, but they can serve as a form of caste-recognition within nests.[172][173] Workers living in monogyne societies tend to be extremely aggressive and attack intruders from neighbouring nests. In queenless colonies, the addition of alien queens or workers does not increase aggression among the population.[174]
Диета

Red imported fire ants are omnivores, and foragers are considered to be қоқыс жинаушылар rather than predators.[175] The ants' diet consists of dead mammals, буынаяқтылар,[176] insects, earthworms, омыртқалылар, and solid food matter such as seeds. However, this species prefers liquid over solid food. The liquid food the ants collect is sweet substances from plants or шіре бал -producing гемиптерандар.[93][175][177][178] Arthropod prey may include dipteran adults, larvae and pupae, and термиттер. The consumption of sugar amino acid is known to affect recruitment of workers to plant nectars. Mimic plants with sugar rarely have workers to feed on them, whereas those with sugar and amino acids have considerable numbers.[179][180] The habitats where they live may determine the food they collect the most; for example, forage success rates for solid foods are highest in lakeshore sites, while high levels of liquid sources were collected from pasture sites.[181] Specific diets can also alter the growth of a colony, with laboratory colonies showing high growth if fed honey-water. Colonies that feed on insects and sugar-water can grow exceptionally large in a short period of time, whereas those that do not feed on sugar-water grow substantially slower. Colonies that do not feed on insects cease brood production entirely.[178] Altogether, the volume of food digested by nestmates is regulated within colonies.[182] Larvae are able to display independent appetites for sources such as solid proteins, amino acid solutions, and sucrose solutions, and they also prefer these sources over dilute solutions. Such behaviour is due to their capability to communicate hunger to workers. The rate of consumption depends on the type, concentration, and state of the food on which they feed. Workers tend to recruit more nestmates to food sources filled with high levels of sucrose than to protein.[182]
Food distribution plays an important role in a colony. This behaviour varies in colonies, with small workers receiving more food than larger workers if a small colony is seriously deprived of food. In larger colonies, however, the larger workers receive more food. Workers can donate sugar water efficiently to other nestmates, with some acting as donors. These "donors" distribute their food sources to recipients, which may also act as donors. Workers may also share a greater portion of their food with other nestmates.[183] In colonies that are not going through starvation, food is still distributed among the workers and larvae. One study shows that honey and soybean oil were fed to the larvae after 12 to 24 hours of being retained by the workers. The ratio distribution of these food sources was 40% towards the larvae and 60% towards the worker for honey, and for soybean oil this figure was around 30 and 70%, respectively.[184] Red imported fire ants also stockpile specific food sources such as insect pieces rather than consuming them immediately. These pieces are usually transported below the mound surface and in the driest and warmest locations.[185]
This species engages in трофаллаксис with the larvae.[186] Regardless of the attributes and conditions of each larva, they are fed roughly the same amount of liquid food. The rate of trophallaxis may increase with larval food deprivation, but such increase depends on the size of each larva. Larvae that are fed regularly tend to be given small amounts. To reach satiation, all larvae regardless of their size generally require the equivalent of eight hours of feeding.[187]
Жыртқыштар

A number of insects, өрмекшітәрізділер, and birds prey on these ants, especially when queens are trying to establish a new colony.[188] While in the absence of defending workers, the fire ant queens must rely on their venom to keep off competitor species [2] Көптеген түрлері инелік, оның ішінде Anax junius, Pachydiplax longipennis, Somatochlora provocans, және Tramea carolina, capture the queens while they are in flight; 16 species of spiders, including the wolf spider Lycosa timuga және southern black widow spider (Latrodectus mactans), actively kill red imported fire ants. L. mactans captures all castes of the species (the workers, queens, and males) within its web. These ants constitute 75% of prey captured by the spider. Кәмелетке толмаған L. mactans spiders have also been seen capturing the ants.[188][189] Other invertebrates that prey on red imported fire ants are earwigs (Labidura riparia ) and tiger beetles (Cicindella punctulata ).[188] Birds that eat these ants include the мұржаның жылдамдығы (Chaetura pelagica), eastern kingbird (Тиранн тиранн), және eastern bobwhite (Colinus virginianus virginianus). The eastern bobwhite attacks these ants by digging out the mounds looking for young queens.[188] Red imported fire ants have been found in stomach contents inside of армадилло.[190]
Many species of ants have been observed attacking queens and killing them. Apparently, the venom of fire ant queens is chemically adapted to rapidly subdue offending competitor ants.[2] Predatory ants include: Ectatomma edentatum, Ephebomyrmex spp., Lasius neoniger, Фейдол spp., Pogonomyrmex badius, және Conomyrma insana, which is among the most significant.[188][191][192] C. insana ants are known to be effective predators against founding queens in studied areas of Northern Florida. The pressure of attacks initiated by C. insana increase over time, causing queens to exhibit different reactions, including escaping, concealment, or defence. Most queens that are attacked by these ants are ultimately killed.[191] Queens that are in groups have higher chances of survival than solitary queens if they are attacked by S. geminata.[193] Ants can attack queens on the ground and invade nests by stinging and dismembering them. Other ants such as P. porcula try to take the head and gaster, and C. clara invade in groups. Also, certain ants try to drag queens out of their nests by pulling on the antennae or legs.[192] Small, monomorphic ants rely on recruitment to kill queens and do not attack them until reinforcements arrive. Aside from killing the queen, some ants may steal the eggs for consumption or emit a repellent that is effective against red imported fire ants.[192] Certain ant species may raid colonies and destroy them.[194]
Parasites, pathogens and viruses

Flies in the genus Псевдактеон (phorid flies) are known to be parasitic to ants. Some species within this genus, such as Pseudacteon tricuspis, have been introduced into the environment for the purpose of controlling the imported fire ant. These flies are parasitoids of the red imported fire ant in its native range in South America, and can be attracted through the ants' venom alkaloids.[195] One species, Pseudacteon obtusus, attacks the ant by landing on the posterioral portion of the head and laying an egg. The location of the egg makes it impossible for the ant to successfully remove it.[196] The larvae migrate to the head, then develop by feeding on the hemolymph, muscle tissue, and nervous tissue. After about two weeks, they cause the ant's head to fall off by releasing an фермент that dissolves the membrane attaching the head to its body. The fly pupates in the detached head capsule, emerging two weeks later.[196][197] P. tricuspis is another phorid fly that is a parasitoid to this species. Although parasitism pressures by these flies do not affect the ants' population density and activity, it has a small effect on a colony population.[198] The стрепсиптеран жәндік Caenocholax fenyesi is known to infect male ants of this species and attack the eggs,[199][200] және кене Pyemotes tritici has been considered a potential biological agent against red imported fire ants, capable of parasitising every caste within the colony.[201] Bacteria, such as Wolbachia, has been found in the red imported fire ant; three different variants of the bacteria are known to infect the red imported fire ant. However, its effect on the ant is unknown.[202][203] Solenopsis daguerrei is a reproductive parasite to red imported fire ant colonies.[204]
A large variety of pathogens and nematodes also infect red imported fire ants. Pathogens include Myrmecomyces annellisae, Маттезия spp., Штайнернема spp.,[205] а мермитид nematode,[206] Vairimorpha invictae, which can be transmitted via live larvae and pupae and dead adults[207][208] және Tetradonema solenopsis, which can be fatal to a large portion of a colony.[209] Individuals infected by Metarhizium anisopliae tend to perform trophallaxis more frequently and have an enhanced preference to хинин, an alkaloid substance.[210] Phorid flies with Kneallhazia solenopsae ретінде қызмет ете алады векторлар in transmitting the disease to the ants.[211] Weakening the colony, infections from this disease are localised within the body fat, with spores only occurring in adult individuals.[212][213] The mortality of an infected colony tends to be greater in contrast to those that are healthy.[206] These ants are a host to Conidiobolus,[214] Myrmicinosporidium durum, және Beauveria bassiana, each of which are parasitic fungi. Infected individuals have spores all over their bodies and appear darker than usual.[215][216][217] The toxicity from antimicrobial property of volatiles produced by the ants can significantly reduce the germination rate of B. bassiana колония ішінде.[218]
A virus, S. invicta 1 (SINV-1 ), has been found in about 20% of fire ant fields, where it appears to cause the slow death of infected colonies. It has proven to be self-sustaining and transmissible. Once introduced, it can eliminate a colony within three months. Researchers believe the virus has potential as a viable биопестицид to control fire ants.[219][220][221] Two more viruses have also been discovered: S. invicta 2 (SINV-2) and S. invicta 3 (SINV-3). Полигинді colonies tend to face greater infections in contrast to monogynous колониялар. Multiple virus infections can also occur.[222][223]
Өмірлік цикл және көбею

Үйлену рейсі in red imported fire ants begins during the warmer seasons of the year (spring and summer), usually two days after rain. Уақыт алатес emerge and mate is between noon and 3:00 кешкі.[224][225] Nuptial flights recorded in North Florida have, on average, 690 female and male alates participating in a single flight.[226] Males are the first to leave the nest, and both sexes readily undertake flight with little to no preflight activity. However, workers swarm the mound excitedly stimulated by mandibular glands within the head of the alates.[227][228] As mounds do not have holes, workers form holes during nuptial flight as a way for the alates to emerge. This behaviour in workers, elicited by the pheromones, includes rapid running and back-and-forth movements, and increased aggression. Workers also cluster themselves around the alates as they climb up on vegetation, and in some cases, attempt to pull them back down before they take flight. Chemical cues from males and females during nuptial flight attract workers, but chemical cues released by workers do not attract other nestmates. It also induces alarm-recruitment behaviour in workers which results in a higher rate of alate retrieval.[227][229]
Males fly at higher elevations than females: captured males are usually 100 to 300 m (330 to 980 ft) above the surface, whereas the females are only 60 to 120 m (200 to 390 ft) above the surface. A nuptial flight takes place for roughly half an hour and females generally fly for less than 1.6 km (0.99 mi) before landing. About 95% of queens successfully mate and only mate once; some males may be infertile due to the testicular lobes failing to develop.[224][230][231][232][233][234] In polygyne colonies, males do not play a significant role and most are, therefore, sterile; one of the reasons for this is to avoid mating with other ant species. This also makes male mortality selective, which may affect the breeding system, mating success and, гендер ағымы.[235][236] Ideal conditions for a nuptial flight to begin is when humidity levels are above 80% and when the soil temperature is above 18 °C (64 °F). Nuptial flights only occur when the ambient temperature is 24–32 °C (75–89 °F).[224]
Queens are often found 1–2.3 miles from the nest they flew from.Colony founding can be done by an individual or in groups, known as pleometrosis.[237] This joint effort of the co-foundresses contributes to the growth and survival of the incipient colony; nests founded by multiple queens begin the growth period with three times as many workers when compared to colonies founded by a single queen. Despite this, such associations are not always stable.[238][239][240][241] The emergence of the first workers instigates queen-queen and queen-worker fighting. In pleometrotic conditions, only one queen emerges victorious, whereas the queens that lost are subsequently killed by the workers.[242] The two factors that could affect the survival of individual queens are their relative fighting capabilities and their relative contribution to worker production. Size, an indicator of fighting capacity, positively correlates with survival rates. However, manipulation of the queen's relative contribution to worker production had no correlation with survival rate.[243]
A single queen lays around 10 to 15 eggs 24 hours after mating.[93] In established nests, a queen applies venom onto each egg that perhaps contains a signal calling for workers to move it.[244] These eggs remain unchanged in size for one week until they hatch into larvae. By this time, the queen will have laid about 75 to 125 more eggs. The larvae that hatch from their eggs are usually covered in their shell membranes for several days. The larvae can free their mouthparts from their shells using body movements, but still need assistance from workers with hatching. The larval stage is divided into four instars, as observed through the moulting stages. At the end of each moult, a piece of unknown material is seen connected to the экзувиялар if they are isolated from the workers. The larval stage lasts between six and 12 days before their bodies expand significantly and become pupae; the pupal stage lasts between nine and 16 days.[44][93]
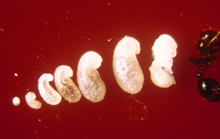
As soon as the first individuals reach the pupal stage, the queen ceases egg production until the first workers mature. This process takes two weeks to one month. The young larvae are fed oils which are regurgitated from her crop, as well as trophic eggs or secretions. She also feeds the young her wing muscles, providing the young with needed nutrients. The first generation of workers are always small because of the limit of nutrients needed for development. These workers are known as minims or nanitics, which burrow out of the queen's chamber and commence foraging for food needed for the colony. Mound construction also occurs at this time. Within a month after the first generation is born, larger workers (major workers) start to develop, and within six months, the mound will be noticeable, if viewed, and houses several thousand residents. A mature queen is capable of laying 1,500 eggs per day; all workers are sterile, so cannot reproduce.[93][245][246][247]
A colony can grow exceptionally fast. Colonies that housed 15–20 workers in May grew to over 7,000 by September. These colonies started to produce reproductive ants when they were a year old, and by the time they were two years old, they had over 25,000 workers. The population doubled to 50,000 when these colonies were three years old.[248] At maturity, a colony can house 100,000 to 250,000 individuals, but other reports suggest that colonies can hold more than 400,000.[d][249][250][251][252] Polygyne colonies have the potential to grow much larger than monogyne colonies.[250]
Several factors contribute to colony growth. Temperature plays a major role in colony growth and development; colony growth ceases below 24 °C and developmental time decreases from 55 days at temperatures of 24 °C to 23 days at 35 °C. Growth in established colonies only occurs at temperatures between 24 and 36 °C. Nanitic brood also develops far quicker than minor worker brood (around 35% faster), which is beneficial for founding colonies.[253] Colonies that have access to an unlimited amount of insect prey are known to grow substantially, but this growth is further accelerated if they are able to access plant resources colonised by гемиптеран жәндіктер.[254] In incipient monogyne colonies where diploid males are produced, colony mortality rates are significantly high and colony growth is slow. In some cases, monogyne colonies experience 100% mortality rates in the early stages of development.[255]

The life expectancy of a worker ant depends on its size, although the overall average is around 62 days.[256] Minor workers are expected to live for about 30 to 60 days, whereas the larger workers live much longer. Larger workers, which have a life expectancy of 60 to 180 days, live 50–140% longer than their smaller counterparts, but this depends on the temperature.[93][257] However, workers kept in laboratory conditions have been known to live for 10 to 70 weeks (70 days to 490 days); the maximum recorded longevity of a worker is 97 weeks (or 679 days).[258] The queens live much longer than the workers, with a lifespan ranging from two years to nearly seven years.[93][258]
In colonies, queens are the only ants able to alter sex ratios which can be predicted. For example, queens originating from male-producing colonies tend to produce predominantly males, while queens that came from female-favoured sex ratio colonies tend to produce females.[259][260] Queens also exert control over the production of sexuals through pheromones that influence the behaviours of workers toward both male and female larvae.[261]
Monogyny and polygyny
There are two forms of society in the red imported fire ant: polygynous colonies and monogynous colonies.[262][263][262] Polygynous colonies differ substantially from monogynous colonies in әлеуметтік жәндіктер. The former experience reductions in queen ұрықтану, dispersal, longevity, and nestmate relatedness.[264][265][266][267] Polygynous queens are also less physogastric than monogynous queens and workers are smaller.[268][269] Understanding the mechanisms behind queen recruitment is integral to understanding how these differences in фитнес қалыптасады It is unusual that the number of older queens in a colony does not influence new queen recruitment. Levels of queen pheromone, which appears to be related to queen number, play important roles in the regulation of reproduction. It would follow that workers would reject new queens when exposed to large quantities of this queen pheromone. Moreover, evidence supports the claim that queens in both populations enter nests at random, without any regard for the number of older queens present.[270] There is no correlation between the number of older queens and the number of newly recruited queens. Three hypotheses have been posited to explain the acceptance of multiple queens into established colonies: мутуализм, туыстық таңдау, және parasitism.[271] The mutualism hypothesis states that cooperation leads to an increase in the personal fitness of older queens. However, this hypothesis is not consistent with the fact that increasing queen number decreases both queen production and queen longevity.[272] Kin selection also seems unlikely given that queens have been observed to cooperate under circumstances where they are statistically unrelated.[273] Сондықтан, патшайымдар жаңа патшайымдарды колонияға жіберу арқылы жеке фитнесінде ешқандай пайда көрмейді. Бұрыннан бар ұялардың паразитизмі полигинияның ең жақсы түсіндірмесі болып көрінеді. Бір теория - колонияға кіруге тырысатындардың көптігі соншалық, жұмысшылар абдырап қалады және байқаусызда оған бірнеше патшайымның қосылуына мүмкіндік береді.[273]
Моногина жұмысшылары шетелдік патшайымдарды өлтіреді және өз аумақтарын агрессивті түрде қорғайды. Алайда, барлық мінез-құлықтар әмбебап бола бермейді, ең алдымен, жұмысшылардың мінез-құлқы олар дамитын экологиялық жағдайға және жұмысшының манипуляциясына байланысты. генотиптер мінез-құлықтың өзгеруін тудыруы мүмкін. Сондықтан жергілікті популяциялардың мінез-құлқы енгізілген популяциялардан өзгеше болуы мүмкін.[274] Моногинді және полигенді қызыл отты құмырсқалар жұмысшыларының агрессивті мінез-құлқын бейтарап ареналардағы өзара әрекеттесуді зерттеу арқылы бағалау және сенімді дамыту этограмма арасында оңай ажыратуға арналған моногине және полигине далада қызыл импортталған өрт құмырсқаларының колониялары,[275] моногинді және полигенді жұмысшылар шыдамдылықтан агрессияға дейінгі әртүрлі мінез-құлықтарымен көрсетілгендей ұя құрбылары мен шетелдіктерді кемсітті. Моногин құмырсқалары әрдайым шетелдік құмырсқаларға, егер олар моногинді немесе полигиндік колониядан болса, дербес шабуыл жасайды, ал полигин құмырсқалары шетелдік полигин құмырсқаларын, негізінен, шабуылдан кейін қалыптасқан мінез-құлыққа ұқсас қалыптармен таныса да, шабуылдамаған. Псевдактеон перидтер. Дұшпандыққа қарсы ескерту мінез-құлқы жұмысшылардың әлеуметтік құрылымына қатты тәуелді болды. Сондықтан шетелдік жұмысшыларға деген мінез-құлық моногинді және полигинді колонияларды сипаттайтын әдіс болды.[276] АҚШ-тың оңтүстік-шығыс және оңтүстік-орталық колонияларының көпшілігі моногинді болып келеді.[241]
Біртекті қызыл импортталған өрт құмырсқасы колониясының аумақтық аумағы мен қорғанның мөлшері оң өзара байланысты, бұл өз кезегінде колония көлемімен (жұмысшылардың саны мен биомассасымен), көршілес колониялардан қашықтығымен, олжалардың тығыздығымен және колонияның ұжымдық бәсекесімен реттеледі. қабілет. Керісінше, полигинді колониялардың ұялы кемсітуі босаңсыған, өйткені қызметкерлер шыдайды ерекше колонияға жат құмырсқалар, басқаларын қабылдайды гетерозигота патшайымдар және өз аумақтарын полигиндік ерекшеліктерден агрессивті түрде қорғамаңыз.[277] Бұл колониялар көптеген патшайымдардың болуы және кооперативті жалдау және бір-бірімен байланысты қорғандар арқылы үлкен аумақтарды пайдалану мүмкіндігінің нәтижесінде репродуктивті өнімді арттыра алады.[278]
Қызыл импортталған өрт құмырсқасында әлеуметтік хромосома бар. Бұл хромосома а-ның екі нұсқасының бірін алып жүретін колонияның әлеуметтік ұйымын ажырата алады суперген (B және b) құрамында 600-ден астам гендер. Әлеуметтік хромосоманы көбінесе жыныстық хромосомалармен салыстырған, өйткені олардың генетикалық ерекшеліктері ұқсас[279] және олар ұқсас жолмен колония фенотипін анықтайды. Мысалы, тек осы хромосоманың В вариантын алып жүретін колониялар жалғыз ВВ патшайымдарын қабылдайды, бірақ В және b нұсқалары бар колониялар тек бірнеше Вб патшайымдарды қабылдайды.[280] Басқа бір геннің айырмашылықтары колонияның жалғыз немесе бірнеше аналық ханшайымға ие болатындығын да анықтай алады.[281]
Басқа организмдермен байланысы

Полигиндер колониялар әлі қалыптаспаған жерлерді басып алған кезде, олардың көптүрлілігі буынаяқтылар және омыртқалылар өте төмендейді.[282] Бұл популяциялар ретінде айқын көрінеді изоподтар, кенелер мен тумбуг тыртықтар айтарлықтай төмендейді. Олар көптеген шыбындар мен қоңыздар тұқымдастарының популяциясын айтарлықтай өзгерте алады, соның ішінде: Calliphoridae, Histeridae, Muscidae, Саркофагида, Silphidae, және Staphylinidae. Осыған қарамастан, бір шолуда кейбір жәндіктерге қызыл импортталған өрт құмырсқалары әсер етпеуі мүмкін екендігі анықталды; мысалы, изоподтардың тығыздығы қызыл импортталған өрт құмырсқалары зақымданған жерлерде азаяды, бірақ крикет тұқымдас Гриллус әсер етпейді. Қызыл жәндіктер мен буынаяқтылар типінің әртүрлілігі көбейетін, қызыл импортталған өрт құмырсқалары кездесетін жерлерде кездесетін жағдайлар бар.[283][284][285] Қызыл импортталған өрт құмырсқалары - үңгір омыртқасыздарының маңызды жыртқыштары, олардың кейбіреулері жойылып бара жатқан түрлер. Бұған кіреді орақшылар, псевдоскорпиондар, өрмекшілер, жер қоңыздары және пселафид қоңыздары. Ең алаңдаушылық - құмырсқаның өзі емес, оларды емдеуге арналған жем, өйткені бұл өлімге әкелуі мүмкін. Сток аралындағы ұлулар (Orthalicus орналасқан) жабайы табиғатта жойылып кетті; импортталған қызыл от құмырсқаларының жыртқыштығы ұлудың жойылуының негізгі факторы болып саналады.[283] Тұтастай алғанда, импортталған қызыл отты құмырсқалар басқаларға қарағанда ерекше буынаяқтыларды жақсы көреді, бірақ олар өздерін қорғай алмайтын немесе қашып кете алмайтын омыртқасыздарға шабуыл жасайды және өлтіреді.[286] Қызыл оттан әкелінетін қызыл құмырсқалар популяциясы азайған немесе жойылғаннан кейін артроподтардың биоәртүрлілігі артады.[114][287]
Қызыл импортталған өрт құмырсқалары мен сүтқоректілердің өзара әрекеттесуі сирек құжатталған. Алайда, тірі аулауда болған жануарлардың қызыл импортталған өрт құмырсқаларының өлімі байқалды.[283][288][289] Өлім деңгейі мақта шығысы (Sylvilagus floridanus) қызыл импортталған өрт құмырсқалары болғандықтан, жастары 33-тен 75% -ке дейін.[290] Қызыл импортталған өрт құмырсқалары көпшілікке қатты әсер етеді деп саналады герпетофауна түрлер; ғалымдар халықтың азаюын атап өтті Флорида патшасы (Lampropeltis getula floridana), және жұмыртқа мен ересектер шығыс қоршау кесірткесі (Sceloporus undulatus) және алты қатарлы жүйрік (Aspidoscelis sexlineata) тамақ көзі болып табылады.[291][292][293] Осыған байланысты шығыс қоршау кесірткелері қызыл импортталған өрт құмырсқасынан құтылу үшін ұзын аяқтары мен жаңа мінез-құлықтарына бейімделді.[294][295] Ересек үш саусақты тасбақалар (Terrapene carolina triunguis), Хьюстон құрбақасы (Anaxyrus houstonensis) кәмелетке толмағандар және Американдық аллигатор (Alligator mississippiensis) балапандар осы құмырсқалар шабуылдап, өлтіреді.[283][296][297][298] Бұл негізінен теріс қауымдастыққа қарамастан, бір зерттеу қызыл импортталған өрт құмырсқалары кене популяциясын реттеу және векторлар мен иелердің динамикасын өзгерту арқылы векторлар арқылы таралатын ауруларға әсер етуі мүмкін екенін көрсетеді, осылайша жануарларға ғана емес, адамдарға да таралу жылдамдығын төмендетеді. .[299]
Өлім-жітімнің деңгейі құстарда жақсы байқалды; өрт құмырсқасының тығыздығы жоғары жерлерде бірде-бір жас ересек адамнан аман қалмаған жағдайлар болды. Көптеген құстар, соның ішінде құзға ұя салатын қарлығаштар, үйрек, аққұтан, бөдене, және терндер қызыл импортталған өрт құмырсқалары әсер етті.[300] Жерге ұя салатын құстар, әсіресе ең аз (Sterna antillarum), құмырсқалардың шабуылына осал.[301][298] Қызыл импортталған өрт құмырсқаларының отарлық асыл тұқымды құстарға әсері әсіресе ауыр; су құстары 100% өлім-жітімге ұшырауы мүмкін, дегенмен бұл фактор ерте ұяға қонатын құстар үшін төмен болды. Аналықтардың тірі қалуы төмендейді Американдық жартас қарлығаштар (Петрохелидон пирронота) егер олар жемшөппен жұмыс жасайтын жұмысшыларға ұшыраса.[302] Songbird қызыл импортталған өрт құмырсқалары бар жерлерде ұяның тірі қалуы азаяды, бірақ тірі қалу деңгейі ақ көзді вирео (Vireo griseus) және қара қақпақты вируэ (Вирео атрикапилла) ұя құмырсқалар жоқ болған кезде немесе оларға шабуыл жасай алмаған кезде 10% -дан 31% -ға және 7% -дан 13% -ға дейін артады.[303] Қызыл импортталған өрт құмырсқалары жанама түрде аналықтардың тірі қалуына ықпал етуі мүмкін Аттоуердің дала тауығы.[304] Алдымен құмырсқалар қыстайтын құстардың азаюымен байланысты деп ойлаған жанжал (Lanius ludovicianus), бірақ кейінірек жүргізілген зерттеу пестицидті қолдана отырып құмырсқаларды жою әрекеттері көрсетті Мирекс, негізінен, улы жанама әсерлері бар екені белгілі болды.[305][306]
Қызыл импортталған өрт құмырсқалары көптеген құмырсқа түрлерімен күшті бәсекелес болып табылады. Олар көптеген табиғи құмырсқаларды ығыстыра алды, бұл бірқатар экологиялық зардаптарға әкелді.[307] Алайда, зерттеулер көрсеткендей, бұл құмырсқалар әрдайым жергілікті құмырсқаларды басатын бәсекелестерден басым бола бермейді. Олар келгенге дейін тіршілік ету ортасын бұзу және жұмысқа қабылдау шектеулері - жергілікті құмырсқалардың басылуының сенімді себептері.[308] Арасында Тапинома меланоцефалия және Феидол фервидасы, қызыл импортталған өрт құмырсқасы екі түрден де күшті, бірақ әртүрлі агрессия деңгейлерін көрсетеді. Мысалы, олар онша жауласпайды T. melanocephalum айырмашылығы P. fervida. Өлім деңгейі T. melanocephalum және P. fervida импортталған қызыл құмырсқалармен күресу жоғары, тиісінше 31,8% және 49,9% құрайды. Қызыл импортталған өрт сөндірушілерінің өлім-жітімі 0,2% -дан 12% -ке дейін.[309] The импортталған жынды құмырсқа (Nylanderia fulvaқызыл импортталған өрт құмырсқасына қарағанда үлкен үстемдік көрсетеді және оларды бір-бірімен кездесетін жерлерде тіршілік ету ортасында ығыстыратыны белгілі.[310] Тротуар құмырсқаларының үлкен колониялары (Tetramorium Caespitum ) қызыл импортталған өрт құмырсқаларының колонияларын жоюы мүмкін, энтомологтар екі түрдің арасындағы бұл қақтығыс қызыл импортталған өрт құмырсқасының таралуына кедергі келтіруі мүмкін деген қорытындыға келді.[311][312] SINV-1 жұқтырған адамдарды сау адамдарға қарағанда тезірек өлтіруге болады Monomorium chinense. Бұл дегеніміз, SINV-1 жұқтырған құмырсқалар сау аналогтарына қарағанда әлсіз және оларды жою мүмкін. M. chinense. Алайда, жұқтырған-жұқтырмағанына қарамастан, негізгі жұмысшылар сирек өлтіріледі.[313]
Қызыл түсті импортталған өрт құмырсқасы әлі күнге дейін доминант болып табылады және 28 құмырсқа түрімен қатар өмір сүреді галерея орманы саңылаулар және он түр ксерофитті орманды шабындық, басқа құмырсқалармен ең агрессивті өзара әрекеттесуді жеңеді. Алайда, кейбір құмырсқалар, мысалы, бірге өмір сүретін жерлерде бірге доминант бола алады Аргентина құмырсқасы, олар симметриялы түрде бәсекелеседі.[314][315] Жұмысшылар басқа құмырсқалармен үнемі тамақ бәсекелестігіне қатысады және бал байыту өндіретін гемиптерандардан (әсіресе Фенакокк соленопсисі ) туған құмырсқалардан; дегенмен қызыл импортталған өрт құмырсқаларын жою мүмкін емес T. melanocephalum толығымен олар тағамның көп бөлігін тұтынады. Керісінше, екі құмырсқа тату-тәтті өмір сүріп, бал арасын бөлісуі мүмкін.[316][317] Көрші өрт құмырсқаларына тап болған кезде, жұмысшылар қабылдауы мүмкін өлім бейнесін жасау оларды сәттілікпен болдырмайтын мінез-құлық. Алайда, мұндай мінез-құлық тек жас жұмысшыларда көрінеді, өйткені жасы үлкен жұмысшылар қашып кетеді немесе қауіп төнген кезде күреседі.[318]
Айтып өткендей, импортталған қызыл құмырсқалар мен аргентиналық құмырсқалар бір-бірімен бәсекелеседі. Өлім коэффициенті әртүрлі сценарийлер бойынша өзгереді (яғни, колониялардың қарсыласуындағы өлім-жітім бір-бірімен далада кездесетіндерден төмен). Ірі жұмысшылар сонымен қатар денелеріндегі көптеген жарақаттарға төтеп бере алады, осылайша аргентиналық құмырсқалардың өлім-жітімін жоғарылатады. Аргентиналық құмырсқалар колониясы 160 000 жұмыскерден тұратын моногинді колонияны ойдағыдай жою үшін колонияға 396 800 жұмысшы қажет болады. Жемді сәтті емдеудің арқасында көлемі кішірейген колония аргентиналық құмырсқалар жыртқыштыққа бейім. Құмырсқалар әлсіреген от құмырсқаларының колонияларын жоюда маңызды рөл атқаруы мүмкін, сонымен қатар бұл өрт құмырсқаларының, әсіресе, аргентиналық құмырсқалар көп шоғырланған жерлерде таралуын бәсеңдетуде маңызды рөл атқаруы мүмкін.[319] Осыған қарамастан, АҚШ-тың оңтүстік-шығысында Аргентинаның құмырсқалар популяциясы қызыл импортталған отты құмырсқа енгізілгеннен кейін азайды.[320]

Қызыл импортталған өрт құмырсқалары инвазиямен қарым-қатынас орнатты тамақ жемісі, Фенакокк соленопсисі. Құмырсқалар өндіретін ресурстарға қол жеткізе алатын болса, колонияның өсуі арта түсетіні белгілі P. solenopsisжәне қызылша әкелінген қызыл құмырсқалар бар жерлерде бұл популяциялардың тығыздығы едәуір жоғары (таралуы) P. solenopsis аудандарда ол құмырсқалардың болуымен байланысты емес). Сонымен қатар, өмір сүру ұзақтығы және репродуктивті коэффициент P. solenopsis екеуі де өседі. Бұл жемшөптерді тіпті жұмысшылар өз ұяларына тасуы мүмкін.[321] Жыртқыштық P. solenopsis табиғи дұшпандардың жыртқыштыққа және паразитизмге кедергі келтіретін жұмысшылардың есебінен азаяды. Асқазанды леди қоңызы аулайды (Menochilus sexmaculatus ) және екі паразиттің иелері болып табылады (Aenasius bambawalei және Acerophagus coccois ) егер құмырсқалар болмаса, бірақ құмырсқалар болса, бұл басқаша. Қызыл қоңыздар қызыл импортталған өрт құмырсқалары бар өсімдіктерде сирек кездеседі, ал «мумия» нимфаларының деңгейі айтарлықтай төмен.[321] Бұл ғалымдар мақта тлиін (Aphis gossypii қызыл импортталған өрт құмырсқасы бар жерлерде популяциялар және күзет құрттары жұмыртқаларының жыртылуы көбейді.[322][323][324] Қызыл импортталған өрт құмырсқалары басқа мылжыңмен өзара қатынасты дамытты (Dysmicoccus morrisoni ). Құмырсқалар колонияның өсуіне ықпал етеді Д.моррисони қорғаныс арқылы колонияларды қоқыстармен жауып, олар шығаратын медовые жинау.[325]
Токсикология
The импортталған қызыл құмырсқа уының токсикологиясы салыстырмалы түрде жақсы зерттелген. The уы құмырсқа үшін маңызды, ол өз олжасын ұстап алуға және өзін қорғауға мүмкіндік береді. Құрама Штаттарда жыл сайын 14 миллионға жуық адам зардап шегеді,[326] олар жеңіл экземадан анафилактикалық шокқа дейінгі реакцияларға ұшырауы мүмкін.[327] Өрттегі құмырсқалардың шағуына реакциялардың көпшілігі - шаншу орнында жану сезімі, содан кейін есекжем және пустула пайда болады.[328] Кейбір зардап шегушілер өмірге қауіп төндіретін аллергиялық реакцияның едәуір асқынуын тудыруы мүмкін.[329][330]У негізінен еритін емес алкалоидтардан тұрады, сулы фазасы бар[331] құрамында 46 ақуыз бар[332] оның төртеуі хабарланды аллергендер.[333][334]
Адамдармен байланыс
Зиянкестер ретінде

Белгілі болуына және инвазиялық мінез-құлқына байланысты қызыл импортталған өрт құмырсқалары қарастырылады зиянкестер. Америка Құрама Штаттарында Азық-түлік және дәрі-дәрмектерді басқару жыл сайын 5 миллиардтан астам доллар емделуге, зақымдануға және зақымданған жерлерде бақылауға жұмсалады.[335][336] Тек Техаста қызыл импортталған өрт құмырсқалары малға, жабайы табиғатқа және халықтың денсаулығына 300 миллион доллар шығын келтірді.[337] Бір американдық үйге шамамен 36 доллардан және қызыл импортталған өрт құмырсқаларын бақылауға және жоюға 250 миллион доллардан астам қаражат жұмсалды. Жеке агенттіктер жыл сайын пестицидтерге 25-40 миллион доллар жұмсайды.[93] Австралиядағы сәтсіз жою экономикаға жыл сайын миллиардтаған шығын әкелуі мүмкін және Квинсленд үкіметінің зерттеуіне сүйенсек, 30 жыл ішінде болжамды шығын 43 миллиард долларға жетуі мүмкін.[338]
Қызыл түсті импортталған өрт құмырсқалары қалалық жерлерде, әсіресе аулаларда, гольф алаңдарында, саябақтарда, демалыс орындарында, мектеп алаңдарында және көше жиектерінде жақсы дамиды, бұл жерде олардың болуы ашық жұмыстарды тоқтатуы мүмкін. Егер олар үйге немесе үйге кірсе, олар үй жануарларына зиян келтіруі мүмкін, егер олар торға салынса, қалам алса, байланған болса немесе қашып құтыла алмаса.[336][339] Ұяларды тротуарлардың, тіпті жолдардың астына, сондай-ақ кіреберістердің, іргетастардың, көгалдардың, тротуарлардың шеттерінің астына, ішкі плитаның астына, электр қораптарына немесе электр желілері маңына салуға болады. Колония топырақты өте көп мөлшерде қаза алады, соның салдарынан кіреберістерде, брусчаткалар мен қабырғаларда құрылымдық проблемалар пайда болады, сонымен қатар жолдарда ойықтардың пайда болуына себеп болуы мүмкін.[93][336][340][341][342] Үйінділерден қосымша зақым ағаштарға, аула өсімдіктеріне және құбырларға келтірілуі мүмкін; кейбір құрылымдар құлап кетуі де мүмкін. Қаныққан топырақты паналау үшін колониялар қатты жаңбырдан кейін адам үйлеріне қоныс аударуы мүмкін.[47][93]
Олар тек қалалық жерлерде ғана дамып қана қоймайды, қызыл импортталған өрт құмырсқалары жабдық пен инфрақұрылымға зиянын тигізіп, бизнеске, жер мен мүлік құндылықтарына әсер етуі мүмкін.[339] Олар сондай-ақ электр қуатына тартылады; электрмен ынталандырылған жұмысшылар уытты алкалоидтарды, дабыл феромондарын шығарады және феромондарды іріктеп алады, бұл өз кезегінде жұмысшыларды алаңға көбірек тартады.[343] Нәтижесінде қызыл импортталған өрт құмырсқалары электр жабдықтарын бұзуы мүмкін.[344] Бұл белгілі магнетизм, мұнда ғалымдар бағдарлау мінез-құлқында рөл атқаратын ішкі магниттік материалдарды анықтады.[345][346] Олар электр оқшаулауын шайнауы белгілі, бұл электр қозғалтқыштарына, суару желілеріне, сорғыларға, сигнал қораптарына, трансформаторларға, телефон станцияларына және басқа жабдықтарға зақым келтіреді.[336][347] Колониялар электр өрістеріне жақын орналасқан және қысқа тұйықталуды тудыруы мүмкін немесе кондиционерлер, компьютерлер және су сорғылары сияқты қосқыштар мен жабдықтарға кедергі келтіруі мүмкін. Олар сондай-ақ әуежайлардың қону алаңдары мен бағдаршамдарды жұқтыратыны белгілі.[347]
Ауыл шаруашылығында

Қызыл импортталған өрт құмырсқасы - бұл жергілікті емес ауылшаруашылық зиянкестері. Олар дақылдарды зақымдауға қабілетті және олар жайылымдарға қауіп төндіреді бақтар.[339] Үйінділердің өзі егін жинау кезінде суару жүйелері сияқты ауылшаруашылық техникасын бұзуы және техниканы зақымдауы мүмкін.[348] Құмырсқалар соя дақылдарын басып кіріп, өнімнің төмен түсуіне әкеліп соқтырады және АҚШ-тың оңтүстік-шығысында соя дақылдары үшін 156 миллион доллар шығын әкелуі мүмкін.[93][349][350] Грузиядан және Солтүстік Каролина 16,8 - 49,1 кг / га сояны өрт құмырсқалары үйінділерінің араласуы салдарынан жинау мүмкін болмады деп мәлімдеу; комбайндар егін жинауға мүмкіндік бермейтін үйінділерден өтіп кетті, ал фермерлер үйінділерге әсер етпеу үшін комбайндарының кескіш штангаларын көтерді.[351][352][353] Қызыл импортталған өрт құмырсқаларының тамақтану тәртібі көптеген басқа дақылдарға, соның ішінде бұршақ, қырыққабат, цитрус, жүгері, қияр, баклажан, окра, жержаңғақ, картоп, құмай, күнбағыс және тәтті картоп. Құмырсқалар сонымен қатар өсімдіктердің тамыр жүйесіне кедергі келтіріп, жас өсіндімен қоректенеді.[93][354] Кейде колониялар цитрус ағаштарының түбіне немесе маңына үйінділер салады, олар жаңа өсімдікті шайнап, гүлдермен немесе дамып келе жатқан жемістермен қоректенеді. Цитрус ағаштары жиі кездеседі белбеу немесе өлтірілді.[355]
Зиянкестердің мәртебесі мен танымал екендігіне қарамастан, қызыл импортталған отты құмырсқа пайдалы болуы мүмкін. Құмырсқа жәндіктердің тиімді жыртқышы болып табылады, сондықтан ол зиянкестердің басқа түрлеріне, әсіресе қант қамысы өрістеріне қарсы биологиялық агент бола алады.[356] Құмырсқа жоятын жәндіктерге мыналар жатады: қоңыздар (Anthonomus grandis) мақта дақылдарында,[357][358][359] қант қамысы бар (Diatraea saccharalis) қант қамысы өрістерінде,[360][361][362] мүйіз шыбыны (Гематобия тітіркендіредікөңде,[363] барқыт құрттары (Anticarsia gemmatalis) сояда, және ақ шыбындар жылыжайларда кездеседі.[364][365] Бірқатар зерттеулер қызыл импортталған өрт құмырсқалары мақта алқаптарында экономикалық маңызы бар жәндіктерге кедергі жасамайтынын және оларға шабуыл жасамайтындығын көрсетеді, соның салдарынан АҚШ-тың оңтүстік-шығыс аймақтарындағы көптеген фермерлер қызыл импортталған өрт құмырсқаларын пайдалы деп санайды.[366] Алайда кейбір ғалымдар география, өсімдік мөлшері, маусымы, топырақтың ылғалдылығы және инсектицидтерді қолдану қарастырылмаған кезде қызыл импортталған өрт құмырсқасының пайдалы мәртебесін болжау қиын деп болжайды. Бұл факторлар зиянкестерге қарсы агент ретінде қызыл импортталған өрт құмырсқаларының тиімділігін төмендетуі мүмкін. Тағы бір фактор - жұмысшылар жайылымдарда мүйіз шыбындары мен басқа зиянкестерді жейтін жыртқыштар мен тли жәндіктер мен масштабты жәндіктердің жыртқыштары сияқты пайдалы жәндіктерді талғамай, жояды.[366][367][368] Олар сонымен қатар құрттары мен қуыршақтарын жеу арқылы паразиттік аралардың зиянкестер түрлеріне қарсы тиімділігін төмендетеді.[366][369]
Бақылау
Сияқты басқа құмырсқа түрлерімен салыстырғанда Anoplolepis gracilipes, олар қамтылған аймақтарды тез басып алатын, қызыл импортталған өрт құмырсқаларын бақылау өте оңай.[370] Құмырсқаны бақылау жөніндегі алғашқы ұсыныстар 1957 жылы пайда болды Америка Құрама Штаттарының конгресі федералдық және штаттық қаржыландыруды қолдана отырып жою бағдарламасын мақұлдады. Құмырсқа және оның биологиясы бойынша зерттеулер жою бағдарламасы құрылғаннан кейін үздіксіз жүргізілді және оларды жою үшін көптеген химиялық заттар қолданылды. Алайда ғалымдар бұл инсектицидтердің табиғи фаунаны өлтіретінін анықтады Қоршаған ортаны қорғау агенттігі кейіннен оларды заңсыз деп тапты. Кейбір ғалымдар құмырсқалардың зиянкестер екендігіне немесе жоқтығына күмән келтірді.[371][372] Бүгінде қызыл шеттен әкелінген отты құмырсқаны АҚШ сияқты аймақтарда жою екіталай. Егер интеграцияланған тәсіл қолданылса, популяцияны дұрыс басқаруға болады.[373] Кейбір ғалымдар құмырсқалардың табиғи дұшпандарын оған қарсы қолдануды ойластырған; бұған кіреді Kneallhazia solenopsae және B. bassiana. Фрид шыбындары потенциалды биологиялық агенттер ретінде қарастырылды, өйткені олар қызыл импортталған өрт құмырсқаларында жемшөп белсенділігін төмендетіп, популяция деңгейіне әсер етуі мүмкін. Алайда олар колонияның өсу жылдамдығына әсер ете алмайды.[374] Сонымен қатар, паразиттік құмырсқалар, паразиттік аралар, кенелер, басқа қоздырғыштар, нематодалар мен саңырауқұлақтар потенциалды биологиялық агенттер болып саналды.[375] Басқалары популяцияны бірнеше экологиялық факторларға манипуляциялау арқылы сақтауға немесе азайтуға болады деп болжайды.[376]

Популяцияны бақылау үшін бірнеше жем қолданылды. Қорғандарға жемдер қолданылса, олар бірнеше апта ішінде жойылады. Суға түсу, шаңдану немесе түтін шығарумен салыстырғанда, жемді қызыл импортталған өрт құмырсқаларына қарсы қолдану тиімді және қарапайым болып саналады. Оларды үйіндіге себеді, содан кейін құмырсқалар алып, оларды жейді.[377][378] Әдетте, колонияны өлтіру үшін өсу реттегіші және бор қышқылы-сахарозаның су жемі, табиғи фаунаның пайдасы және төмен концентрация сияқты жемдер қажет.[379][380] Қызыл импортталған өрт құмырсқаларына қарсы қолданылатын басқаларға жем жатады Амдро, Көтерілу, гидраметилнон және Maxforce.[381]
Қатты және сұйық жемді инсектицидтер, егер бұл жерде дұрыс қолданылмаса, су ағыны арқылы қажетсіз су көздеріне жіберілуі мүмкін. Көбінесе, бұл көптеген диффузиялық көздер, соның ішінде нүктелік емес көздер арқылы болады. Қар еріп, жауын-шашын жер үстімен және үстімен жылжып келе жатқанда, су дұрыс қолданылмаған инсектицидтерді жинап алып, оларды үлкенірек су қоймаларына, өзендерге, батпақты жерлерге немесе су қоймаларына перколаттарға жинайды.
Зерттеушілер сонымен қатар фумигация, үйіндіге ине салу, үйіндіге су құю және жер үсті шаңын тазарту сияқты басқа әдістермен тәжірибе жүргізіп келеді. Үйіндіден суды ағызу қорғанға улы сұйықтықтың көп мөлшерін құюға әкеп соғады. Бұл қорғанның көп бөлігіне әсер етуі мүмкін болса да, ол патшайымға жетпеуі мүмкін, осылайша колонияның бұзылуына жол бермейді. Беттік шаңдану үйінді суды ағызуға ұқсас, тек инсектицидтер үйінді үстіне жағылады және дымқыл болған кезде топыраққа сіңеді. Үйінді инъекциясында қысым мен инсектицидтерді үйіндіге енгізу қолданылады, бірақ патшайымға бұл әсер етпеуі мүмкін. Үйінділерді фумигациялау арқылы тиімді түрде жоюға болады. Трансляция арқылы емдеу кейде ауылшаруашылық құрал-жабдықтарымен зарарланған аймақтарды бүрку арқылы қолданылады. Сонымен, инсектицидтермен спотты өңдеуді үйіндіге инсектицидтің қалдықтарын бұрғылау және енгізу арқылы қолдануға болады.[93][377]
Химиялық бақылаудан басқа, осы құмырсқаларға қарсы қолданылатын басқа әдістерге механикалық және электрлік құрылғылар жатады. Алайда, бұл құрылғылардың тиімді екендігі белгісіз. Құмырсқалардан қорғану ғимараттардың ішіне ұя салатын, жарықтарды бітеу және бітеу жолымен тиімді болуы мүмкін, бұл қабырғалардан тыс жерлердегі тұрғындарды сәтті басады. Үй иелері үйінділерді қайнаған су құйып немесе тез тұтанатын сұйықтықтармен тұтандырып, жоюдың өзіндік әдістерін қолданды. Бұл әдістер тиімді болғанымен, олар ұсынылмайды, өйткені олар адамдарға және қоршаған ортаға зиян тигізуі мүмкін.[93][377]
Ескертулер
- ^ Дегенмен S. geminata кладограммада пайда болады, ол түр тобына кірмейді. Ол тек «тіркеме нүктесі» ретінде қызмет ету үшін енгізілген; дәлірек айтсақ, бұл тексеріліп отырған кладограмманың қай жермен байланысып жатқанын көрсетеді өмір ағашы.[28]
- ^ Кейбір деректер олар 1929 жылы келген деп болжайды.[104]
- ^ Қызыл импортталған өрт құмырсқаларына жету ықтимал Жаңа Орлеан жылы Луизиана, сонымен қатар, бірақ Аргентиналық құмырсқалар сол жерде құмырсқалардың қоныстануына және таралуына жол бермеді.[105]
- ^ Бір колонияның халқы маусымдық жағынан әр түрлі болуы мүмкін.[249]
Әдебиеттер тізімі
Сілтемелер
- ^ Джонсон, Н.Ф. (19 желтоқсан 2007). "Solenopsis invicta Бөрен «. Hymenoptera Name Server 1.5 нұсқасы. Колумбус, Огайо, АҚШ: Огайо мемлекеттік университеті. Архивтелген түпнұсқа 2016 жылғы 20 қыркүйекте. Алынған 1 сәуір 2015.
- ^ а б в Фокс, Эдуардо Г.П .; Ву, Сяоцин; Ван, Лей; Чен, Ли; Лу, Ён-Юэ; Сюй, Цзыюань (ақпан 2019). «Изозоленопсин патшайымның уы құмырсқа бәсекелестерінің жылдам әрекет қабілеттілігін төмендетеді». Токсикон. 158: 77–83. дои:10.1016 / j.toxicon.2018.11.428. PMID 30529381. S2CID 54481057.
- ^ Tschinkel 2006, б. vii.
- ^ Taber 2000, б. 12.
- ^ Боуерсок, Г.В .; Браун, П. (1999). Кеш антикалық кезең: постклассикалық әлемге нұсқаулық (2-ші басылым). Кембридж: Гарвард университетінің Belknap баспасы. б.284. ISBN 978-0-674-51173-6.
- ^ Старр, С .; Эверс, С .; Старр, Л. (2010). Биология: Физиологиясыз түсініктер мен қосымшалар (8-ші басылым). Белмонт, Калифорния: Cengage Learning. б.431. ISBN 978-0-538-73925-2.
- ^ Льюис, П.Х. (1990 жылғы 24 шілде). «Оңтүстіктен шыққан қуатты от құмырсқалары». The New York Times. Алынған 2 қараша 2016.
- ^ Қоршаған орта, су, мұра және өнер бөлімі. «Түрлер банкі: Solenopsis invicta (Formicidae отбасы) «. Австралиялық биологиялық ресурстарды зерттеу: Австралиялық фауналар каталогы. Канберра: Австралия үкіметі. Архивтелген түпнұсқа 21 қыркүйек 2016 ж. Алынған 8 сәуір 2014.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ Tschinkel 2006, 13-14 бет.
- ^ Дризес, Б.М. (2002). «Қызыл импортталған өрт құмырсқасының медициналық мәселелері және емдеу мәселелері» (PDF). Техас импортталған өрт қауырсындарын зерттеу және басқару жобасы. Texas A&M University. Алынған 23 тамыз 2016.
- ^ Бут, К .; Дами, М. (2008). Қызыл импортталған өрт құмырсқасы (Solenopsis invicta): Колония жасын анықтауға қатысты әдебиеттерге шолу (PDF). Биоалуантүрлілік және биоқауіпсіздік орталығы (Есеп). Бастапқы өнеркәсіп министрлігі: биологиялық қауіпсіздік Жаңа Зеландия. б. 2. мұрағатталған түпнұсқа (PDF) 2016 жылғы 9 қазанда. Алынған 29 қараша 2018.
- ^ Мастерсон, Дж. (2007). «Түрлердің атауы: Solenopsis invicta". Смитсон теңіз станциясы. Архивтелген түпнұсқа 2016 жылғы 20 қыркүйекте. Алынған 2 сәуір 2016.
- ^ Кармайкл, А. (2005). «Қызыл импортталған өрт құмырсқасы Solenopsis invicta Бурен (Hymenoptera: Formicidae: Myrmicinae) «. PaDIL (Австралиялық биоқауіпсіздік). Алынған 2 сәуір 2016.
- ^ Бухс 2005 ж, б. 13.
- ^ а б Санцки, Ф. (1916). «Formicides sudaméricains nouveaux ou peu connus». Физ. 2: 365–399. дои:10.5281 / ZENODO.14374.
- ^ Taber 2000, б. 25.
- ^ а б в Шаттук, С.О .; Портер, С.Д .; W., D.P. (1999). «3069 іс. Solenopsis invicta Бюрен, 1972 (Insecta, Hymenoptera): нақты атауды сақтау ұсынылған ». Зоологиялық номенклатура бюллетені. 56 (1): 27–30. дои:10.5962 / bhl.3030-бөлім. ISSN 0007-5167.
- ^ Крейтон, В.С. (1930). «Тұқымның жаңа әлемдік түрлері Соленопсис (Hymenop. Formicidae) « (PDF). Американдық өнер және ғылым академиясының еңбектері. 66 (2): 39–152. дои:10.2307/20026320. JSTOR 20026320. Архивтелген түпнұсқа (PDF) 2016 жылғы 4 наурызда.
- ^ Уилсон, Э.О. (1952). «The Solenopsis saevissima Оңтүстік Америкадағы кешен (Hymenoptera: Formicidae) ». Memórias do Instituto Oswaldo Cruz. 50: 60–68. дои:10.1590 / S0074-02761952000100003. ISSN 0074-0276. PMID 13012835.
- ^ Этершанк, Г. (1966). «Мирмицина әлеміне қатысты жалпы ревизия Соленопсис және Федологетон (Hymenoptera: Formicidae) ». Австралия зоология журналы. 14 (1): 73–171. дои:10.1071 / ZO9660073.
- ^ Кемпф, В.В. (1972). «Каталог abreviado das formigas da Regiao Neotropical». Studia Entomologica (N.S.). 15: 3–344.
- ^ а б в г. e f ж сағ мен j к л м n o Бурен, В.Ф. (1972). «Импортталған өрт құмырсқаларының таксономиясы бойынша ревизиялық зерттеулер». Джорджия энтомологиялық қоғамының журналы. 7: 1–26. дои:10.5281 / zenodo.27055.
- ^ а б в г. Trager, JC (1991). «От құмырсқаларын қайта қарау, Solenopsis geminata топ (Hymenoptera: Formicidae: Myrmicinae) «. Нью-Йорк энтомологиялық қоғамының журналы. 99 (2): 141–198. дои:10.5281 / zenodo.24912. JSTOR 25009890.
- ^ а б Болтон, Б. (1995). Әлемдегі құмырсқалардың жаңа жалпы каталогы. Кембридж: Гарвард университетінің баспасы. 388-391 бет. ISBN 978-0-674-61514-4.
- ^ Taber 2000, б. 26.
- ^ Халықаралық зоологиялық номенклатура жөніндегі комиссия (2001). «Пікір 1976 ж Solenopsis invicta Buren, 1972 (Insecta, Hymenoptera): нақты атауы сақталған «. Зоологиялық номенклатура бюллетені. 58 (2): 156–157. ISSN 0007-5167.
- ^ Болтон, Б. (2016). "Solenopsis invicta". AntCat. Алынған 19 тамыз 2016.
- ^ а б Tschinkel 2006, б. 14.
- ^ а б Росс, К.Г .; Meer, RKV; Флетчер, Дж .; Варго, Э.Л. (1987). «Екі енгізілген отты құмырсқалар мен олардың гибридтерін биохимиялық фенотиптік және генетикалық зерттеу (Hymenoptera: Formicidae)». Эволюция. 41 (2): 280–293. дои:10.2307/2409138. JSTOR 2409138. PMID 28568766.
- ^ Росс, К.Г .; Варго, Э.Л .; Флетчер, Дэвид Дж. (1987). «Солтүстік Америкадағы үш отты құмырсқа түрінің салыстырмалы биохимиялық генетикасы, екі әлеуметтік формасына ерекше сілтеме жасай отырып Solenopsis invicta (Hymenoptera: Formicidae) ». Эволюция. 41 (5): 979–990. дои:10.2307/2409186. JSTOR 2409186. PMID 28563420.
- ^ Tschinkel 2006, б. 15–16.
- ^ а б Питтс, Дж .; McHugh, JV .; Росс, К.Г. (2005). «Өрт құмырсқаларын кладистикалық талдау Solenopsis saevissima түр-топ (Hymenoptera: Formicidae) ». Zoologica Scripta. 34 (5): 493–505. дои:10.1111 / j.1463-6409.2005.00203.x.
- ^ Tschinkel 2006, б. 503.
- ^ Питтс, Дж.П. (2002). Solenopsis saevissima түр-тобына кладистикалық талдау (Hymenoptera: Formicidae) (PDF) (Ph.D.). Джорджия университеті, Афина. 42-43 бет.
- ^ Taber 2000, б. 43.
- ^ Росс, К.Г .; Етікші, Д.Д. (1997). «От құмырсқасының екі әлеуметтік формасындағы ядролық және митохондриялық генетикалық құрылым Solenopsis invicta: балама әлеуметтік ұйымға өту туралы түсініктер ». Тұқымқуалаушылық. 78 (6): 590–602. дои:10.1038 / hdy.1997.98. S2CID 19225329.
- ^ Кригер, МДж .; Росс, К.Г .; Чанг, С..В.Й .; Келлер, Л. (1999). «От құмырсқасындағы триплоидияның жиілігі мен шығу тегі Solenopsis invicta". Тұқымқуалаушылық. 82 (2): 142–150. дои:10.1038 / sj.hdy.6884600. S2CID 13144037.
- ^ Келлер, Л .; Росс, К.Г. (1998). «Өзімшіл гендер: қызыл отты құмырсқадағы жасыл сақал» (PDF). Табиғат. 394 (6693): 573–575. Бибкод:1998 ж.394..573K. дои:10.1038/29064. S2CID 4310467.
- ^ Графен, А. (1998). «Эволюциялық биология: жасыл сақал өлім туралы бұйрық» (PDF). Табиғат. 394 (6693): 521–522. Бибкод:1998 ж.394..521G. дои:10.1038/28948. S2CID 28124873.
- ^ Вюрм, Ю .; Ванг Дж .; Риба-Грогнуз, О .; Корона, М .; Ньюгард, С .; Хант, Б.Г .; Инграм, К.К .; Фолкет, Л .; Нипитваттанафон, М .; Готзек, Д .; Дайкстра, М.Б .; Оеттлер, Дж .; Комтесс Ф .; Ших, С.-Дж .; Ву, В.-Дж .; Янг, С-С .; Томас, Дж .; Бодинг, Э .; Прадерванд, С .; Флегель, V .; Кук, Э.Д .; Фаббретти, Р .; Стокингер, Х .; Ұзын, Л .; Фармери, В.Г .; Оуки Дж .; Бумсма, Дж. Дж .; Памило, П .; Ии, С.В .; Хайнце, Дж .; Гудисман, М.А .; Фаринелли, Л .; Харшман, К .; Хуло, Н .; Церутти, Л .; Ксенариос, Мен .; Етікші Д .; Келлер, Л. (2011). «От құмырсқасының геномы Solenopsis invicta" (PDF). Ұлттық ғылым академиясының материалдары. 108 (14): 5679–5684. Бибкод:2011PNAS..108.5679W. дои:10.1073 / pnas.1009690108. PMC 3078418. PMID 21282665.
- ^ Хеджер, SA (1997). Мореланд, Д. (ред.) Зиянкестермен күресу жөніндегі анықтама (8-ші басылым). Mallis анықтамалығы және техникалық оқыту компаниясы. 531-535 бб.
- ^ Tschinkel 2006, б. 24.
- ^ Бухс 2005 ж, б. 11.
- ^ а б в Петралия, Р.С .; Винсон, С.Б (1979). «Импортталған от құмырсқасының дернәсілдері мен жұмыртқаларының даму морфологиясы, Solenopsis invicta". Америка энтомологиялық қоғамының жылнамалары. 72 (4): 472–484. дои:10.1093 / aesa / 72.4.472.
- ^ а б Уилер, Г.С .; Уилер, Дж. (1977). «Құмырсқа дернәсілдеріне қосымша зерттеулер: Myrmicinae». Американдық энтомологиялық қоғамның операциялары. 103: 581–602. дои:10.5281 / zenodo.25100.
- ^ Петралия, Р.С .; Соренсен, А.А .; Винсон, С.Б. (1980). «Импортталған өрт құмырсқасының дернәсілдерінің лабия безі жүйесі, Solenopsis invicta Бөрен ». Жасушалар мен тіндерді зерттеу. 206 (1): 145–156. дои:10.1007 / BF00233615. PMID 6153574. S2CID 28786982.
- ^ а б Гринберг, Л .; Кабашима, Дж.Н. (2014). «Зиянкестер туралы ескертулер: Қызыл импортталған өрт құмырсқасы». Мемлекеттік зиянкестермен күресудің интеграцияланған бағдарламасы. Ауыл шаруашылығы және табиғи ресурстар, Калифорния университеті. Алынған 4 сәуір 2016.
- ^ О'Нил, Дж .; Маркин, Г.П. (1975). «Импортталған от құмырсқасының личинкалық шағылыстары, Solenopsis invicta Бурен (Hymenoptera: Formicidae) ». Канзас энтомологиялық қоғамының журналы. 48 (2): 141–151. JSTOR 25082731.
- ^ а б в г. Фокс, Эдуардо Г П; Солис, Даниэль Рус; Росси, Моника Ланзони; Делаби, Жак Губерт Шарль; де Соуза, Родриго Фернандо; Буэно, Одаир Корреа (2012). «Бразилиялық отты құмырсқалардың салыстырмалы жетілмеген морфологиясы (Hymenoptera: Formicidae: Solenopsis)». Психика: Энтомология журналы. 2012: 1–10. дои:10.1155/2012/183284.
- ^ Миренда, Дж. Т .; Винсон, С.Б. (1981). «Еңбек бөлінісі және қызыл импортталған өрт құмырсқасындағы касталардың спецификациясы Solenopsis invicta Бөрен ». Жануарлардың мінез-құлқы. 29 (2): 410–420. дои:10.1016 / S0003-3472 (81) 80100-5. S2CID 53199246.
- ^ Schmid-Hempel 1998 ж, б. 144.
- ^ Hölldobler & Wilson 1990, б. 311.
- ^ Hölldobler & Wilson 1990, б. 318.
- ^ а б Цчинкель, В.Р. (1988). «Колонияның өсуі және от құмырсқасындағы жұмысшы полиморфизмінің онтогенезі, Solenopsis invicta". Мінез-құлық экологиясы және социобиология. 22 (2): 103–115. дои:10.1007 / BF00303545. S2CID 8455126.
- ^ а б в г. e Цчинкель, В.Р .; Михеев, А.С .; Storz, S.R. (2003). «От құмырсқасы жұмысшыларының аллометриясы, Solenopsis invicta". Жәндіктер туралы журнал. 3 (2): 2. дои:10.1673/031.003.0201. PMC 524642. PMID 15841219.
- ^ Портер, С.Д .; Цчинкель, В.Р. (1985). «Өрт құмырсқасының полиморфизмі: асыл тұқымды эргономика». Мінез-құлық экологиясы және социобиология. 16 (4): 323–336. дои:10.1007 / BF00295545. JSTOR 4599785. S2CID 5967997.
- ^ а б Фогт, Дж .; Аппел, А.Г. (2000). «Өрт құмырсқасындағы үздіксіз газ алмасу, Solenopsis invicta Бурен: Касталық айырмашылықтар және температураның әсерлері ». Жәндіктер физиологиясы журналы. 46 (4): 403–416. дои:10.1016 / S0022-1910 (99) 00123-7. PMID 12770204.
- ^ а б Клоуден, МЖ (2007). Жәндіктердегі физиологиялық жүйелер. Амстердам; Бостон: Elsevier / Academic Press. 357–383, 433–449 бб. ISBN 978-0-12-369493-5.
- ^ Хилл, Р.В .; Вайс, Г.А .; Андерсон, М. (2012). Жануарлар физиологиясы (3-ші басылым). Сандерленд, Массачусетс: Sinauer Associates. 612-614 бет. ISBN 978-0-87893-662-5.
- ^ а б в г. e f Винсон, С.Б. (1983). «Импортталған өрт құмырсқасының физиологиясы қайта қаралды». Флорида энтомологы. 66 (1): 126–139. дои:10.2307/3494559. JSTOR 3494559.
- ^ Петралия, Р.С .; Уильямс, Х.Дж .; Винсон, С.Б. (1982). «Артқы ішектің ультрақұрылымы және импортталған өрт құмырсқасының дернәсілдерінің шығаратын өнімдері,» Solenopsis invicta Бөрен ». Sociaux жәндіктері. 29 (2): 332–345. дои:10.1007 / BF02228760. S2CID 7324976.
- ^ Флетчер, Дж .; Блум, М.С. (1981). «Патшайым өрт құмырсқаларындағы дигиляцияны және оогенезді феромоналды бақылау». Ғылым. 212 (4490): 73–75. Бибкод:1981Sci ... 212 ... 73F. дои:10.1126 / ғылым.212.4490.73. PMID 17747633.
- ^ Баркер, Дж.Ф. (1978). «Импортталған өрт құмырсқасындағы ооциттердің жетілуін нейроэндокриндік реттеу Solenopsis invicta". Жалпы және салыстырмалы эндокринология. 35 (3): 234–237. дои:10.1016/0016-6480(78)90067-9. PMID 689357.
- ^ Джонс, Р.Г .; Дэвис, У.Л .; Хунг, А.К.Ф .; Винсон, С.Б. (1978). «Өрт құмырсқаларындағы ұшу бұлшықетінің ұрықтандырудан туындаған гистолизі (Соленопсис, спп.): ультрақұрылымдық зерттеу ». Американдық анатомия журналы. 151 (4): 603–610. дои:10.1002 / aja.1001510411. PMID 645619.
- ^ Том, П.М .; Джонсон, К.П .; Cupp, E.W. (1976). «Дене резервтерін алдын-ала эксплуатациялау кезінде пайдалану Solenopsis invicta". Америка энтомологиялық қоғамының жылнамалары. 69 (1): 145–148. дои:10.1093 / aesa / 69.1.145.
- ^ а б Винсон, С.Б .; Филлипс, С.А .; Уильямс, Х.Ж. (1980). «Қызыл импортталған өрт құмырсқасының жұтқыншақтан кейінгі бездерінің қызметі, Solenopsis invicta buren ». Жәндіктер физиологиясы журналы. 26 (9): 645–650. дои:10.1016/0022-1910(80)90035-9.
- ^ Вандер Меер, Р.К .; Лофгрен, С.С. (1989). «Өрт құмырсқалары арасындағы дұшпанның биохимиялық және мінез-құлықты дәлелдеуі, Solenopsis invicta және Solenopsis richteri (Hymenoptera: Formicidae) ». Химиялық экология журналы. 15 (6): 1757–1765. дои:10.1007 / BF01012263. PMID 24272179. S2CID 23144401.
- ^ а б Вандер Меер, Р.К .; Глэнси, Б.М .; Лофгрен, СС .; Гловер, А .; Тумлинсон, Дж. Х .; Рокка, Дж. (1980). «Қызыл импортталған өрт құмырсқасының патшайымдарының улы қабы: феромонды тартудың көзі». Америка энтомологиялық қоғамының жылнамалары. 73 (5): 609–612. дои:10.1093 / aesa / 73.5.609.
- ^ Чой, М .; Вандер Меер, Р.К .; Рену, М. (2012). «Құмырсқа ізі феромонды биосинтез нейропептидті гормонмен қоздырылады». PLOS ONE. 7 (11): e50400. Бибкод:2012PLoSO ... 750400C. дои:10.1371 / journal.pone.0050400. PMC 3511524. PMID 23226278.
- ^ Elzen, GW (1986). «Импортталған өрт құмырсқасындағы оттегі шығыны және судың жоғалуы Solenopsis invicta Бөрен ». Салыстырмалы биохимия және физиология А. 84 (1): 13–17. дои:10.1016/0300-9629(86)90035-6.
- ^ Чен Дж .; Рашид Т .; Фэн, Г .; Хьюз, В. (2014). «Арасындағы салыстырмалы зерттеу Solenopsis invicta және Solenopsis richteri ыстыққа және десекция стрессіне төзімділік туралы ». PLOS ONE. 9 (6): e96842. Бибкод:2014PLoSO ... 996842C. дои:10.1371 / journal.pone.0096842. PMC 4051589. PMID 24915009.
- ^ Бойлз, Дж .; Обри, Д.П .; Хикман, Кр .; Мюррей, К.Л .; Тимпон, Дж .; Ops, C.H. (2009). «Қызыл импортталған өрт құмырсқаларының физиологиялық реакциясының өзгеруі (Solenopsis invicta) шағын масштабты жылулық біртектілікке дейін ». Термиялық биология журналы. 34 (2): 81–84. дои:10.1016 / j.jtherbio.2008.10.005.
- ^ а б Портер, С.Д .; Цчинкель, В.Р. (1993). «Өрт құмырсқасының термиялық артықшылықтары: өсу мен метаболизмнің мінез-құлқын бақылау» Мінез-құлық экологиясы және социобиология. 32 (5). дои:10.1007 / BF00183787. S2CID 9840395.
- ^ а б Фогт, Дж .; Appel, AG (1999). «От құмырсқасының стандартты метаболикалық жылдамдығы, Solenopsis invicta Бурен: температураның, массаның және кастаның әсері ». Жәндіктер физиологиясы журналы. 45 (7): 655–666. дои:10.1016 / S0022-1910 (99) 00036-0. PMID 12770351.
- ^ а б в г. e f ж сағ мен j к Веттерер, Дж. (2013). «Экзотикалық таралуы Solenopsis invicta (Hymenoptera: Formicidae) Солтүстік Америкадан тыс ». Әлеуметтану. 60: 50–55. дои:10.13102 / социобиология.v60i1.50-55.
- ^ а б Аллен, Г. Е .; Бурен, В.Ф .; Уильямс, Р. Н .; Менезес, М. Д .; Whitcomb, W. H. (1974). «Қызыл импортталған өрт құмырсқасы, Solenopsis invicta; Mato Grosso, Бразилиядағы таралуы және тіршілік ету ортасы ». Америка энтомологиялық қоғамының жылнамалары. 67 (1): 43–46. дои:10.1093 / aesa / 67.1.43.
- ^ а б Мешер, МС .; Росс, К.Г .; Етікші, Д.Д .; Келлер, Л .; Krieger, MJ B. (2003). «От құмырсқасының екі әлеуметтік формасының таралуы Solenopsis invicta (Hymenoptera: Formicidae) туған жері Оңтүстік Америкада « (PDF). Америка энтомологиялық қоғамының жылнамалары. 96 (6): 810–817. дои:10.1603 / 0013-8746 (2003) 096 [0810: DOTTSF] 2.0.CO; 2.[тұрақты өлі сілтеме ]
- ^ а б Noordijk, J. (2010). «Нидерландыдағы өрт құмырсқаларына қауіп-қатерді талдау». Лейден: Еуропалық омыртқасыздарды зерттеу. б. 15. Алынған 5 сәуір 2016.
- ^ а б в г. e Бурен, В.Ф .; Аллен, Дж .; Уиткомб, В.Х .; Леннартц, Ф.Е .; Уильямс, Р.Н. (1974). «Импортталған өрт құмырсқаларының зоогеографиясы». Нью-Йорк энтомологиялық қоғамының журналы. 82 (2): 113–124. JSTOR 25008914.
- ^ Кальдера, Э.Дж .; Росс, К.Г .; DeHeer, ChJ .; Етікші, Д.Д .. (2008). «Инвазиялық от құмырсқасының туынды көзі Solenopsis invicta АҚШ-та »деп аталады. Биологиялық инвазиялар. 10 (8): 1457–1479. дои:10.1007 / s10530-008-9219-0. S2CID 4471306.
- ^ Пескуэро, М.А .; Dias, AMP (2011). «Географиялық өтпелі аймақ Соленопсис өрт құмырсқалары (Hymenoptera: Formicidae) және Псевдактеон Сан-Паулу штатында паразитоидтер (Diptera: Phoridae) ұшады, Бразилия ». Неотропикалық энтомология. 40 (6): 647–652. дои:10.1590 / S1519-566X2011000600003. PMID 23939270.
- ^ а б в г. «Түрлер: Solenopsis invicta Buren, 1972". AntWeb. Калифорния ғылым академиясы. Алынған 5 сәуір 2016.
- ^ Wild, A.L. (2007). "A catalogue of the ants of Paraguay (Hymenoptera: Formicidae)" (PDF). Зоотакса. 1622: 1–55. дои:10.11646/zootaxa.1622.1.1. ISSN 1175-5334.
- ^ Santschi, F. (1923). «Соленопсис et autres fourmis néotropicales". Revue Suisse de Zoologie. 30 (8): 245–273. дои:10.5281/ZENODO.14217.
- ^ Ahrens, M.E.; Ross, K.G.; Shoemaker, D.D. (2005). "Phylogeographic structure of the fire ant Solenopsis invicta in its native South American range: roles of natural barriers and habitat connectivity" (PDF). Эволюция. 59 (8): 1733–1743. дои:10.1111/j.0014-3820.2005.tb01822.x. PMID 16329243.[тұрақты өлі сілтеме ]
- ^ а б в г. e f "Solenopsis invicta (red imported fire ant)". Инвазивті түрлер жиынтығы. CABI. 2014 жыл. Алынған 12 сәуір 2016.
- ^ Taber 2000, б. 28.
- ^ Cokendolpher, J.C.; Phillips, S.A. Jr. (1989). "Rate of spread of the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae), in Texas". Оңтүстік-Батыс натуралисті. 34 (3): 443–449. дои:10.2307/3672182. JSTOR 3672182.
- ^ "Solenopsis invicta (insect)". Инвазивті түрлердің ғаламдық базасы. Invasive Species Specialist Group. 2010 жыл. Алынған 2 сәуір 2016.
- ^ Buhs 2005, б. 12.
- ^ а б в г. e Kemp, S.F.; deShazo, R.D.; Moffitt, J.E.; Williams, D.F.; Buhner, W.A. (2000). "Expanding habitat of the imported fire ant (Solenopsis invicta): a public health concern". Аллергия және клиникалық иммунология журналы. 105 (4): 683–691. дои:10.1067/mai.2000.105707. PMID 10756216. S2CID 8280947.
- ^ Desforges, J.F.; deShazo, R.D.; Butcher, B.T.; Banks, W.A. (1990). "Reactions to the stings of the imported fire ant". Жаңа Англия Медицина журналы. 323 (7): 462–466. дои:10.1056/NEJM199008163230707. PMID 2197555.
- ^ а б в г. e f ж сағ мен j к л м n o Collins, L.; Scheffrahn, R.H. (2001). "Red imported fire ant, Solenopsis invicta Buren". UF/IFAS Featured Creatures. University of Florida. Архивтелген түпнұсқа on 21 September 2016. Алынған 13 сәуір 2016.
- ^ "Ant Colonies and Social Structure | Terminix". www.terminix.com. Алынған 19 қараша 2019.
- ^ Hubbard, M.D.; Cunningham, W.G. (1977). "Orientation of mounds in the ant Solenopsis invicta (Hymenoptera, Formicidae, Myrmicinae)". Sociaux жәндіктері. 24 (1): 3–7. дои:10.1007/BF02223276. S2CID 5851856.
- ^ Penick, C.A.; Tschinkel, W. R. (2008). "Thermoregulatory brood transport in the fire ant, Solenopsis invicta". Sociaux жәндіктері. 55 (2): 176–182. дои:10.1007/s00040-008-0987-4. S2CID 15284230.
- ^ Cassill, D.; Tschinkel, W.R.; Vinson, S.B. (2002). "Nest complexity, group size and brood rearing in the fire ant, Solenopsis invicta". Sociaux жәндіктері. 49 (2): 158–163. дои:10.1007/s00040-002-8296-9. hdl:10806/3290. S2CID 13938249.
- ^ Taber 2000, б. 29.
- ^ Taber 2000, б. 31.
- ^ "100 of the World's Worst Invasive Alien Species". Инвазивті түрлердің ғаламдық базасы. Invasive Species Specialist Group. Алынған 26 қаңтар 2014.
- ^ Mooallem, J. (5 December 2013). "There's a Reason They Call Them 'Crazy Ants'". The New York Times. Алынған 23 тамыз 2016.
- ^ Fountain, H. (8 December 2008). "Fire Ants Win Out Through Land Changes, Not a Better Build". The New York Times. Алынған 31 қазан 2016.
- ^ King, J.R.; Tschinkel, W.R. (2008). "Experimental evidence that human impacts drive fire ant invasions and ecological change". Ұлттық ғылым академиясының материалдары. 105 (51): 20339–20343. Бибкод:2008PNAS..10520339K. дои:10.1073/pnas.0809423105. PMC 2629336. PMID 19064909.
- ^ Capinera 2008, б. 3116.
- ^ Buhs 2005, б. 20.
- ^ Wilcove, D.S.; Rothstein, D.; Dubow, J.; Филлипс, А .; Losos, E. (1998). "Quantifying threats to imperiled species in the United States" (PDF). BioScience. 48 (8): 607–615. дои:10.2307/1313420. JSTOR 1313420. Архивтелген түпнұсқа (PDF) 2011 жылғы 12 тамызда.
- ^ Tschinkel 2006, б. 28.
- ^ Buhs 2005, б. 9.
- ^ Fountain, H. (8 July 2008). "Tracing an Ant Invasion to a Handful of Queens". The New York Times. Алынған 2 қараша 2016.
- ^ Ross, K.G; Shoemaker, D.D. (2008). "Estimation of the number of founders of an invasive pest insect population: the fire ant Solenopsis invicta in the USA". Корольдік қоғамның еңбектері B: Биологиялық ғылымдар. 275 (1648): 2231–2240. дои:10.1098/rspb.2008.0412. PMC 2603238. PMID 18577505.
- ^ Tschinkel 2006, б. 25.
- ^ Buhs 2005, б. 23.
- ^ Adkins, H.G. (1970). "The imported fire ant in the Southern United States". Америкалық географтар қауымдастығының жылнамалары. 60 (3): 578–592. дои:10.1111/j.1467-8306.1970.tb00742.x.
- ^ а б Epperson, D.M.; Allen, C.R. (2010). "Red imported fire ant impacts on upland arthropods in southern Mississippi". The American Midland Naturalist. 163 (1): 54–63. дои:10.1674/0003-0031-163.1.54. S2CID 12992133.
- ^ Callcott, A.-M.; Collins, H.L. (1996). "Invasion and range expansion of imported fire ants (Hymenoptera: Formicidae) in North America from 1918–1995". Флорида энтомологы. 79 (2): 240–251. дои:10.2307/3495821. JSTOR 3495821.
- ^ Korzukhin, M.D.; Porter, S.D.; Thompson, L.C.; Wiley, S. (2001). "Modeling temperature-dependent range limits for the fire ant Solenopsis invicta (Hymenoptera: Formicidae) in the United States" (PDF). Environmental Entomology. 30 (4): 645–655. дои:10.1603/0046-225X-30.4.645. S2CID 53065271.
- ^ Callcott, A.-M.; Oi, D.H.; Collins, H.L.; Williams, D.F.; Lockley, T.C. (2000). "Seasonal studies of an isolated red imported fire ant (Hymenoptera: Formicidae) population in eastern Tennessee" (PDF). Environmental Entomology. 29 (4): 788–794. дои:10.1603/0046-225X-29.4.788. S2CID 53073021.
- ^ Pimm, S.L.; Bartell, D.P. (1980). "Statistical model for predicting range expansion of the red imported fire ant, Solenopsis invicta, in Texas". Environmental Entomology. 9 (5): 653–658. дои:10.1093/ee/9.5.653.
- ^ Capinera 2008, pp. 2034–2035.
- ^ Morrison, L.W.; Korzukhin, M.D.; Porter, S.D. (2005). "Predicted range expansion of the invasive fire ant, Solenopsis invicta, in the eastern United States based on the VEMAP global warming scenario". Әртүрлілік және таралуы. 11 (3): 199–204. дои:10.1111/j.1366-9516.2005.00142.x. S2CID 46089176.
- ^ а б Morrison, L.W.; Porter, S.D.; Daniels, E.; Korzukhin, M.D. (2004). "Potential global range expansion of the invasive fire ant, Solenopsis invicta" (PDF). Biological Invasions. 6 (2): 183–191. дои:10.1023/B:BINV.0000022135.96042.90. S2CID 9973667.[тұрақты өлі сілтеме ]
- ^ Buhs 2005, б. 5.
- ^ "Red imported fire ant - Solenopsis invicta". Қоршаған ортаны қорғау департаменті. Government of Australia. Архивтелген түпнұсқа on 21 September 2016. Алынған 21 наурыз 2015.
- ^ McCubbin, K.I.; Weiner, J.M. (2002). "Fire ants in Australia: a new medical and ecological hazard". Австралияның медициналық журналы. 176 (11): 518–519. дои:10.5694/j.1326-5377.2002.tb04547.x. PMID 12064981.
- ^ Henshaw, Michael T; Kunzmann, N.; Vanderwoude, C.; Sanetra, M.; Crozier, R.H (2005). "Population genetics and history of the introduced fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae), in Australia". Австралиялық энтомология журналы. 44 (1): 37–44. дои:10.1111/j.1440-6055.2005.00421.x.
- ^ Condon, M. (27 July 2013). "Queensland launched a war against the fire ant invasion, but 12 years later, they're still on the march". Курьерлік пошта. Алынған 30 желтоқсан 2014.
- ^ Willis, P. (22 February 2007). "Fire ant update". ABC News. Архивтелген түпнұсқа on 21 September 2016. Алынған 4 мамыр 2015.
- ^ "National fire ant eradication program". Бастапқы өндіріс бөлімі. Квинсленд үкіметі. 2007. мұрағатталған түпнұсқа on 21 October 2009.
- ^ "Importation of Red Imported Fire Ants Solenopsis invicta Buren 1972 - profile". Қоршаған орта және мұра бөлімі. Government of NSW. 2014. мұрағатталған түпнұсқа on 21 September 2016. Алынған 5 мамыр 2015.
- ^ Creedon, K. (4 December 2014). "Race against time to quarantine Sydney outbreak of red fire ants". 9 жаңалықтар. Сидней, Австралия. Архивтелген түпнұсқа on 21 September 2016. Алынған 6 желтоқсан 2014.
- ^ Australian Associated Press (25 February 2016). "US fire ants found at Brisbane airport". SBS News. Архивтелген түпнұсқа on 21 September 2016. Алынған 23 тамыз 2016.
- ^ Moloney, S.; Vanderwoude, C. (2002). "Red imported fire ants: A threat to eastern Australia's wildlife?". Экологиялық менеджмент және қалпына келтіру. 3 (3): 167–175. дои:10.1046/j.1442-8903.2002.t01-1-00109.x. S2CID 54926916.
- ^ Rajagopal, T.; Sevarkodiyone, S.P.; Sekar, M. (2005). "Ant species richness, diversity and similarity index at five selected localities of Sattur Taluk". Indian Journal of Environmental Education. 5: 7–12.
- ^ Na, J.P.S.; Ли, C.Y. (2001). "Identification key to common urban pest ants in Malaysia" (PDF). Tropical Biomedicine. 18 (1): 1–17.
- ^ Kuo, K.C. (2008). Management of Red Invasive Fire Ants and Fruit Flies: The Taiwan Experience (PDF). Taipei, Taiwan: Food & Fertilizer Technology Center (FFTC). б. 1.
- ^ Бхарти, Х .; Guénard, B.; Bharti, M.; Economo, E.P. (2016). "An updated checklist of the ants of India with their specific distributions in Indian states (Hymenoptera, Formicidae)". ZooKeys (551): 1–83. дои:10.3897/zookeys.551.6767. PMC 4741291. PMID 26877665.
- ^ Tschinkel 2006, б. 72.
- ^ Ванг, Л .; Yongyue, L.; Yijuan, X.; Ling, Z. (2013). "The current status of research on Solenopsis invicta Buren (Hymenoptera: Formicidae) in mainland China" (PDF). Asian Myrmecology. 5: 125–137. ISSN 1985-1944.
- ^ Zeng, L.; Lu, Y.Y.; He, X.F.; Zhang, W.Q.; Liang, G.W. (2005). "Identification of red imported fire ant Solenopsis invicta to invade mainland China and infestation in Wuchuan, Guangdong". Chinese Bulletin of Entomology. 42 (2): 144–148. ISSN 0452-8255.
- ^ Чжан, Р .; Ли, Ю .; Лю, Н .; Porter, S.D. (2007). "An overview of the red imported fire ant (Hymenoptera: Formicidae) in Mainland China". Флорида энтомологы. 90 (4): 723–731. дои:10.1653/0015-4040(2007)90[723:aootri]2.0.co;2. Алынған 12 сәуір 2016.
- ^ Sutherst, R.W.; Maywald, G. (2005). "A climate model of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae): implications for invasion of new regions, particularly Oceania". Environmental Entomology. 34 (2): 317–335. дои:10.1603/0046-225X-34.2.317. S2CID 85951260.
- ^ Buren, W.F. (1982). "Red imported fire ant now in Puerto Rico". Флорида энтомологы. 65 (1): 188–189. дои:10.2307/3494163. JSTOR 3494163.
- ^ Wetterer, J.K.; Snelling, R.R. (2006). "The red imported fire ant, Solenopsis invicta, in the Virgin Islands (Hymenoptera: Formicidae)" (PDF). Флорида энтомологы. 89 (4): 431–434. дои:10.1653/0015-4040(2006)89[431:TRIFAS]2.0.CO;2.
- ^ Deyrup, M.; Davis, L.; Buckner, S. (1998). Composition of the Ant Fauna of Three Bahamian Islands (PDF). San Salvador: Proceedings of the 7th Symposium on the Natural History of the Bahamas. б. 27. ISBN 978-0-935909-66-1.
- ^ Davis, L.R.; Vander Meer, R.K.; Porter, S.D. (2001). "Red imported fire ants expand their range across the West Indies" (PDF). Флорида энтомологы. 84 (4): 735. дои:10.2307/3496416. JSTOR 3496416.
- ^ Ascunce, M.S.; Yang, C.-C.; Oakey, J.; Calcaterra, L.; Wu, W.-J.; Shih, C.-J.; Goudet, J.; Ross, K.G.; Shoemaker, D. (2011). "Global invasion history of the fire ant Solenopsis invicta". Ғылым. 331 (6020): 1066–1068. Бибкод:2011Sci...331.1066A. дои:10.1126/science.1198734. PMID 21350177. S2CID 28149214.
- ^ Mlot, N.J.; Craig, A.T.; Hu, D.L. (2011). "Fire ants self-assemble into waterproof rafts to survive floods". Ұлттық ғылым академиясының материалдары. 108 (19): 7669–7673. Бибкод:2011PNAS..108.7669M. дои:10.1073/pnas.1016658108. PMC 3093451. PMID 21518911.
- ^ Morrill, W.L. (1974). "Dispersal of red imported fire ants by water". Флорида энтомологы. 57 (1): 39–42. дои:10.2307/3493830. JSTOR 3493830.
- ^ Hölldobler & Wilson 1990, б. 171.
- ^ а б Adams, B.J.; Hooper-Bùi, L.M.; Strecker, R.M.; O'Brien, D.M. (2011). "Raft formation by the red imported fire ant, Solenopsis invicta". Жәндіктер туралы журнал. 11 (171): 171. дои:10.1673/031.011.17101. PMC 3462402. PMID 22950473.
- ^ Haight, K.L. (2006). "Defensiveness of the fire ant, Solenopsis invicta, is increased during colony rafting". Sociaux жәндіктері. 53 (1): 32–36. дои:10.1007/s00040-005-0832-y. S2CID 24420242.
- ^ Howard, D.F.; Tschinkel, W.R. (1976). "Aspects of necrophoric behavior in the red imported fire ant, Solenopsis invicta" (PDF). Мінез-құлық. 56 (1): 157–178. дои:10.1163/156853976x00334. ISSN 0005-7959.
- ^ Qiu, H.-L.; Lu, L.-H.; Shi, Q.-X.; Tu, C.-C.; Lin, T.; He, Y.-R. (2015). "Differential necrophoric behaviour of the ant Solenopsis invicta towards fungal-infected corpses of workers and pupae". Энтомологиялық зерттеулер жаршысы. 105 (5): 607–614. дои:10.1017/S0007485315000528. PMID 26082426.
- ^ Smith, C.W.; Frederiksen, R.A. (2000). Sorghum: Origin, History, Technology, and Production. New York, New York: John Wiley & Sons. б. 456. ISBN 978-0-471-24237-6.
- ^ Zettler, J.A.; Spira, T.P.; Allen, C.R. (2001). "Ant–seed mutualisms: can red imported fire ants sour the relationship?". Биологиялық сақтау. 101 (2): 249–253. дои:10.1016/S0006-3207(01)00074-X.
- ^ Cumberland, M.S.; Kirkman, L.K. (2013). "The effects of the red imported fire ant on seed fate in the longleaf pine ecosystem". Өсімдіктер экологиясы. 214 (5): 717–724. дои:10.1007/s11258-013-0201-2. S2CID 2648579.
- ^ Markin, G.P.; O'Neal, J.; Dillier, J. (1975). "Foraging tunnels of the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae) ». Канзас энтомологиялық қоғамының журналы. 48 (1): 83–89. JSTOR 25082717.
- ^ а б в г. Porter, S.D.; Tschinkel, W.R. (1987). "Foraging in Solenopsis invicta (Hymenoptera: Formicidae): effects of weather and season". Environmental Entomology. 16 (3): 802–808. дои:10.1093/ee/16.3.802.
- ^ Showler, A.T.; Knaus, R.M.; Reagan, T.E. (1989). "Foraging territoriality of the imported fire ant, Solenopsis invicta Buren, in surgarcane as determined by neutron activation analysis". Sociaux жәндіктері. 36 (3): 235–239. дои:10.1007/BF02226306. S2CID 36832531.
- ^ Гравиш, Н .; Monaenkova, D.; Goodisman, M.A.D.; Goldman, D.I. (2013). "Climbing, falling, and jamming during ant locomotion in confined environments". Ұлттық ғылым академиясының материалдары. 110 (24): 9746–9751. arXiv:1305.5860. Бибкод:2013PNAS..110.9746G. дои:10.1073/pnas.1302428110. PMC 3683784. PMID 23690589.
- ^ а б Yijuan, X.; Yongyue, L.; Линг, З .; Guangwen, L. (2007). "Foraging behavior and recruitment of red imported fire ant Solenopsis invicta Buren in typical habitats of South China". Acta Ecologica Sinica. 27 (3): 855–860. дои:10.1016/S1872-2032(07)60022-5.
- ^ Tschinkel, W.R. (2011). "The organization of foraging in the fire ant, Solenopsis invicta". Жәндіктер туралы журнал. 11 (26): 26. дои:10.1673/031.011.0126. PMC 3391925. PMID 21529150.
- ^ а б в г. Vander Meer, R.K. (1983). "Semiochemicals and the red imported fire ant (Solenopsis invicta Buren) (Hymenoptera: Formicidae)". Флорида энтомологы. 66 (1): 139–161. дои:10.2307/3494560. JSTOR 3494560.
- ^ Williams, H.J.; Strand, M.R.; Vinson, S.B. (1981). "Trail pheromone of the red imported fire ant Solenopsis invicta (Buren)". Experientia. 37 (11): 1159–1160. дои:10.1007/BF01989893. S2CID 46572483.
- ^ Vander Meer, R.K.; Alvarez, F.; Lofgren, C.S. (1988). "Isolation of the trail recruitment pheromone of Solenopsis invicta". Химиялық экология журналы. 14 (3): 825–838. дои:10.1007/BF01018776. PMID 24276134. S2CID 32388563.
- ^ Barlin, M.R.; Блум, М.С .; Brand, M.M. (1976). "Fire ant trail pheromones: Analysis of species specificity after gas chromatographic fractionation". Journal of Insect Physiology. 22 (6): 839–844. дои:10.1016/0022-1910(76)90253-5.
- ^ Capinera 2008, б. 3445.
- ^ Taber 2000, б. 41.
- ^ Vander Meer, R.K.; Preston, C.A.; Choi, M.-Y. (2010). "Isolation of a pyrazine alarm pheromone component from the fire ant, Solenopsis invicta". Химиялық экология журналы. 36 (2): 163–170. дои:10.1007/s10886-010-9743-0. PMID 20145982. S2CID 10622781.
- ^ а б Hubbard, M.D. (1974). "Influence of nest material and colony odor on digging in the ant Solenopsis invicta (Hymenoptera: Formicidae) ». Georgia Entomological Society. 9: 127–132.
- ^ Obin, Martin S. (1986). "Nestmate recognition cues in laboratory and field colonies of Solenopsis invicta Buren (Hymenoptera: Formicidae)". Химиялық экология журналы. 12 (9): 1965–1975. дои:10.1007/BF01041858. PMID 24305971. S2CID 19517580.
- ^ Obin, M.S.; Vander Meer, R.K. (1989). "Nestmate recognition in fire ants (Solenopsis invicta Buren). Do queens label workers?". Этология. 80 (1–4): 255–264. дои:10.1111/j.1439-0310.1989.tb00744.x.
- ^ Vander Meer, R.K.; Preston, C.A.; Hefetz, A. (2008). "Queen regulates biogenic amine level and nestmate recognition in workers of the fire ant, Solenopsis invicta". Naturwissenschaften. 95 (12): 1155–1158. Бибкод:2008NW.....95.1155V. дои:10.1007/s00114-008-0432-6. PMID 18704354. S2CID 7640153.
- ^ Obin, M.S.; Vander Meer, R.K. (1988). "Sources of nestmate recognition cues in the imported fire ant Solenopsis invicta Buren (Hymenoptera: Formicidae)". Жануарлардың мінез-құлқы. 36 (5): 1361–1370. дои:10.1016/S0003-3472(88)80205-7. S2CID 53205306.
- ^ а б Taber 2000, б. 37.
- ^ Taber 2000, б. 38.
- ^ Tennant, L.E.; Porter, S.D. (1991). "Comparison of diets of two fire ant species (Hymenoptera: Formicidae): solid and liquid components". Journal of Entomological Science. 26 (4): 450–465. дои:10.18474/0749-8004-26.4.450. ISSN 0749-8004.
- ^ а б Porter, S.D. (1989). "Effects of diet on the growth of laboratory fire ant colonies (Hymenoptera: Formicidae)". Канзас энтомологиялық қоғамының журналы. 62 (2): 288–291. JSTOR 25085088.
- ^ Lanza, J.; Vargo, E. L.; Pulim, S.; Chang, Y. Z. (1993). "Preferences of the fire ants Solenopsis invicta және S. geminata (Hymenoptera: Formicidae) for amino acid and sugar components of extrafloral nectars". Environmental Entomology. 22 (2): 411–417. дои:10.1093/ee/22.2.411. S2CID 17682096.
- ^ Lanza, J. (1991). "Response of fire ants (Formicidae: Solenopsis invicta және S.gerninata) to artificial nectars with amino acids". Экологиялық энтомология. 16 (2): 203–210. дои:10.1111/j.1365-2311.1991.tb00210.x.
- ^ Vogt, J.T.; Grantham, R.A.; Corbett, E.; Rice, S.A.; Wright, R.E. (2002). "Dietary habits of Solenopsis invicta (Hymenoptera: Formicidae) in four Oklahoma habitats". Environmental Entomology. 31 (1): 47–53. дои:10.1603/0046-225X-31.1.47. S2CID 85586177.
- ^ а б Cassill, D.L.; Tschinkel, W.R. (1999). "Regulation of diet in the fire ant, Solenopsis invicta" (PDF). Жәндіктердің мінез-құлық журналы. 12 (3): 307–328. дои:10.1023/A:1020835304713. S2CID 18581840.
- ^ Howard, D.F.; Tschinkel, W.R. (1980). "The effect of colony size and starvation on food flow in the fire ant, Solenopsis invicta (Hymenoptera: Formicidae) ». Мінез-құлық экологиясы және социобиология. 7 (4): 293–300. дои:10.1007/BF00300670. JSTOR 4599344. S2CID 18507664.
- ^ Sorensen, A.A.; Vinson, S.B. (1981). "Quantitative food distribution studies within laboratory colonies of the imported fire ant, Solenopsis invicta Buren". Sociaux жәндіктері. 28 (2): 129–160. дои:10.1007/BF02223701. S2CID 28896246.
- ^ Gayahan, G.G.; Tschinkel, W.R. (2008). "Fire ants, Solenopsis invicta, dry and store insect pieces for later use". Жәндіктер туралы журнал. 8 (39): 39. дои:10.1673/031.008.3901. PMC 3127378.
- ^ Taber 2000, б. 39.
- ^ Cassill, D.L.; Tschinkel, W.R. (1995). "Allocation of liquid food to larvae via trophallaxis in colonies of the fire ant, Solenopsis invicta". Жануарлардың мінез-құлқы. 50 (3): 801–813. дои:10.1016/0003-3472(95)80140-5. S2CID 18395869.
- ^ а б в г. e Whitcomb, W.H.; Bhatkar, A.; Nickerson, J.C. (1973). "Predators of Solenopsis invicta queens prior to successful colony establishment". Environmental Entomology. 2 (6): 1101–1103. дои:10.1093/ee/2.6.1101.
- ^ Nyffeler, M.; Dean, D.A.; Sterling, W.L. (1988). "The southern black widow spider, Latrodectus mactans (Araneae, Theridiidae), as a predator of the red imported fire ant, Solenopsis invicta (Hymenoptera, Formicidae), in Texas cotton fields". Journal of Applied Entomology. 106 (1–5): 52–57. дои:10.1111/j.1439-0418.1988.tb00563.x.
- ^ Deyrup, M.; Trager, J. (1986). "Ants of the Archbold Biological Station, Highlands County, Florida (Hymenoptera: Formicidae)". Флорида энтомологы. 69 (1): 206–228. дои:10.2307/3494763. JSTOR 3494763.
- ^ а б Nickerson, J.C.; Whitcomb, W.H.; Bhatkar, A.P.; Naves, M.A. (1975). "Predation on founding queens of Solenopsis invicta by workers of Conomyrma insana". Флорида энтомологы. 58 (2): 75–82. дои:10.2307/3493384. JSTOR 3493384.
- ^ а б в Nichols, B.J.; Sites, R.W. (1991). "Ant predators of founder queens of Solenopsis invicta (Hymenoptera: Formicidae) in central Texas". Environmental Entomology. 20 (4): 1024–1029. дои:10.1093/ee/20.4.1024.
- ^ Jerome, C.A.; McInnes, D.A.; Adams, E.S. (1998). "Group defense by colony-founding queens in the fire ant Solenopsis invicta". Мінез-құлық экологиясы. 9 (3): 301–308. дои:10.1093/beheco/9.3.301.
- ^ Rao, Asha; Vinson, S. B. (2004). "Ability of resident ants to destruct small colonies of Solenopsis invicta (Hymenoptera: Formicidae) ». Environmental Entomology. 33 (3): 587–598. дои:10.1603/0046-225X-33.3.587. S2CID 85657533.
- ^ Чен, Ли; Sharma, K.R.; Fadamiro, H.Y. (2009). "Fire ant venom alkaloids act as key attractants for the parasitic phorid fly, Pseudacteon tricuspis (Diptera: Phoridae)". Naturwissenschaften. 96 (12): 1421–1429. Бибкод:2009NW.....96.1421C. дои:10.1007/s00114-009-0598-6. PMID 19714317. S2CID 24205012.
- ^ а б Williams, D.F.; Banks, W.A. (1987). «Pseudacteon obtusus (Diptera: Phoridae) attacking Solenopsis invicta (Hymenoptera: Formicidae) in Brazil". Psyche: A Journal of Entomology. 94 (1–2): 9–13. дои:10.1155/1987/85157.
- ^ Ehrenberg, R. (18 September 2009). "Venom attracts decapitating flies". Ғылым жаңалықтары. Архивтелген түпнұсқа 2016 жылғы 22 қыркүйекте. Алынған 30 желтоқсан 2014.(жазылу қажет)
- ^ Morrison, L.W.; Porter, S.D. (2005). "Testing for population-level impacts of introduced Pseudacteon tricuspis flies, phorid parasitoids of Solenopsis invicta fire ants". Биологиялық бақылау. 33 (1): 9–19. CiteSeerX 10.1.1.542.4377. дои:10.1016/j.biocontrol.2005.01.004.
- ^ Schmid-Hempel 1998, б. 94.
- ^ Катиритамби, Дж .; Johnston, J.S. (1992). "Stylopization of Solenopsis invicta (Hymenoptera: Formicidae) by Caenocholax fenyesi (Strepsiptera: Myrmecolacidae) in Texas". Америка энтомологиялық қоғамының жылнамалары. 85 (3): 293–297. дои:10.1093/aesa/85.3.293.
- ^ Bruce, W.A.; LeCato, G.L. (1980). «Pyemotes tritici: A potential new agent for biological control of the red imported fire ant, Solenopsis invicta (Acari: Pyemotidae)". International Journal of Acarology. 6 (4): 271–274. дои:10.1080/01647958008683230.
- ^ Ahrens, M.E.; Shoemaker, D. (2005). "Evolutionary history of Wolbachia infections in the fire ant Solenopsis invicta". BMC эволюциялық биологиясы. 5: 35. дои:10.1186/1471-2148-5-35. PMC 1175846. PMID 15927071.
- ^ Schmid-Hempel 1998, б. 34.
- ^ Capinera 2008, б. 188.
- ^ Schmid-Hempel 1998, б. 59.
- ^ а б Briano, J.; Calcaterra, L.; Varone, L. (2012). "Fire ants (Соленопсис spp.) and their natural enemies in southern South America". Psyche: A Journal of Entomology. 2012: 1–19. дои:10.1155/2012/198084.
- ^ Jouvenaz, D.P.; Ellis, E.A. (1986). «Vairimorpha invictae N. Sp. (Microspora: Microsporida), a parasite of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae)". Протозоология журналы. 33 (4): 457–461. дои:10.1111/j.1550-7408.1986.tb05641.x.
- ^ Oi, D.H.; Briano, J. A.; Valles, S.M.; Williams, D.F. (2005). "Transmission of Vairimorpha invictae (Microsporidia: Burenellidae) infections between red imported fire ant (Hymenoptera: Formicidae) colonies". Омыртқасыздар патологиясы журналы. 88 (2): 108–115. дои:10.1016/j.jip.2004.11.006. PMID 15766927.
- ^ Nickle, W.R.; Jouvenaz, D.P. (1987). "Tetradonema solenopsis n. sp. (Nematoda: Tetradonematidae) parasitic on the red imported fire ant Solenopsis invicta Buren from Brazil". Journal of Nematology. 19 (3): 311–313. PMC 2618658. PMID 19290149.
- ^ Qiu, H.-L.; Lu, L.-H.; Zalucki, M.-P.; He, Y.-R. (2016). "Metarhizium anisopliae infection alters feeding and trophallactic behavior in the ant Solenopsis invicta" (PDF). Омыртқасыздар патологиясы журналы. 138: 24–29. дои:10.1016/j.jip.2016.05.005. PMID 27234423.
- ^ Durham, S. (2010). "ARS Parasite Collections Assist Research and Diagnoses". Ауылшаруашылық ғылыми-зерттеу қызметі. Америка Құрама Штаттарының Ауыл шаруашылығы министрлігі. Алынған 25 тамыз 2014.
- ^ Knell, J.D.; Allen, G.E.; Hazard, E.I. (1977). "Light and electron microscope study of Thelohania solenopsae n. sp. (Microsporida: Protozoa) in the red imported fire ant, Solenopsis invicta". Омыртқасыздар патологиясы журналы. 29 (2): 192–200. дои:10.1016/0022-2011(77)90193-8. PMID 850074.
- ^ "South fights fire ants with pathogen". CNN. 1 маусым 1998 ж. Алынған 1 қараша 2016.
- ^ Schmid-Hempel 1998, б. 295.
- ^ Pereira, R.M. (2004). «Пайда болуы Myrmicinosporidium durum in red imported fire ant, Solenopsis invicta, and other new host ants in eastern United States". Омыртқасыздар патологиясы журналы. 86 (1–2): 38–44. дои:10.1016/j.jip.2004.03.005. PMID 15145249.
- ^ Schmid-Hempel 1998, б. 44.
- ^ Tschinkel 2006, б. 632.
- ^ Ванг, Л .; Эллиотт, Б .; Джин, Х .; Zeng, L.; Chen, J. (2015). "Antimicrobial properties of nest volatiles in red imported fire ants, Solenopsis invicta (hymenoptera: formicidae)". Naturwissenschaften. 102 (11–12): 66. Бибкод:2015SciNa.102...66W. дои:10.1007/s00114-015-1316-1. PMID 26467352. S2CID 1650319.
- ^ Valles, S.M.; Strong, C.A.; Dang, P.M.; Hunter, W.B.; Pereira, R.M.; Oi, D.H.; Shapiro, A.M.; Williams, D.F. (2004). "A picorna-like virus from the red imported fire ant, Solenopsis invicta: initial discovery, genome sequence, and characterization" (PDF). Вирусология. 328 (1): 151–157. дои:10.1016/j.virol.2004.07.016. PMID 15380366.
- ^ Valles, S.M.; Strong, C.A.; Oi, D.H.; Porter, S.D.; Pereira, R.M.; Vander Meer, R.K.; Хашимото, Ю .; Hooper-Bùi, L.M.; Sánchez-Arroyo, H.; Davis, T.; Karpakakunjaram, V.; Vail, K.M.; Fudd Graham, L.C.; Briano, J.A.; Calcaterra, L.A.; Gilbert, L.E.; Ward, R.; Уорд, К .; Oliver, J.B.; Taniguchi, G.; Thompson, D.C. (2007). "Phenology, distribution, and host specificity of Solenopsis invicta virus-1". Омыртқасыздар патологиясы журналы. 96 (1): 18–27. дои:10.1016/j.jip.2007.02.006. PMID 17412359.
- ^ "Fire ants may have met their match". CNN. Associated Press. 7 May 2007. Archived from түпнұсқа on 13 May 2007.
- ^ Valles, S.M.; Hashimoto, Y. (2009). «Оқшаулау және сипаттама Solenopsis invicta virus 3, a new positive-strand RNA virus infecting the red imported fire ant, Solenopsis invicta". Вирусология. 388 (2): 354–61. дои:10.1016/j.virol.2009.03.028. PMID 19403154.
- ^ Аллен, С .; Valles, S.M.; Күшті, C.A. (2011). "Multiple virus infections occur in individual polygyne and monogyne Solenopsis invicta ants". Омыртқасыздар патологиясы журналы. 107 (2): 107–111. дои:10.1016/j.jip.2011.03.005. PMID 21439294.
- ^ а б в Trager 1988, б. 419.
- ^ Buhs 2005, б. 21.
- ^ Morrill, W.L. (1974). "Production and flight of alate red imported fire ants". Environmental Entomology. 3 (2): 265–271. дои:10.1093/ee/3.2.265.
- ^ а б Alonso, L.E.; Vander Meer, R.K. (1997). "Source of alate excitant pheromones in the red imported fire ant Solenopsis invicta (Hymenoptera: Formicidae) ». Жәндіктердің мінез-құлық журналы. 10 (4): 541–555. дои:10.1007/BF02765376. S2CID 12295884.
- ^ Markin, G.P.; Dillier, J.H.; Hill, S.O.; Блум, М.С .; Hermann, H.R. (1971). "Nuptial flight and flight ranges of the imported fire ant, Solenopsis saevissima richteri (Hymenoptera: Formicidae) ». Journal of the Georgia Entomological Society. 6 (3): 145–156.
- ^ Obin, M.S.; Vander Meer, R.K. (1994). "Alate semiochemicals release worker behavior during fire ant nuptial flights" (PDF). Journal of Entomological Science. 29 (1): 143–151. дои:10.18474/0749-8004-29.1.143.
- ^ Hung, A.C.F.; Vinson, S. B.; Summerlin, J. W. (1974). "Male sterility in the red imported fire ant, Solenopsis invicta". Америка энтомологиялық қоғамының жылнамалары. 67 (6): 909–912. дои:10.1093/aesa/67.6.909.
- ^ Trager 1988, б. 422.
- ^ Capinera 2008, б. 3444.
- ^ Capinera 2008, б. 3112.
- ^ Hölldobler & Wilson 1990, б. 156.
- ^ Fritz, G.N.; Vander Meer, R.K.; Preston, C.A. (2006). "Selective male mortality in the red imported fire ant, Solenopsis invicta". Генетика. 173 (1): 207–213. дои:10.1534/genetics.106.056358. PMC 1461442. PMID 16489215.
- ^ Hölldobler & Wilson 1990, б. 145.
- ^ Tschinkel, W. (1992). "Brood raiding and the population dynamics of founding and incipient colonies of the fire ant, Solenopsis invicta". Экологиялық энтомология. 17 (2): 179–188. дои:10.1111/j.1365-2311.1992.tb01176.x.
- ^ Goodisman, M.; Ross, K. (1999). "Queen recruitment in a multiple-queen population of the fire ant Solenopsis invicta". Мінез-құлық экологиясы. 10 (4): 428–435. дои:10.1093/beheco/10.4.428.
- ^ Tschinkel, W.R.; Howard, D.F. (1983). "Colony founding by pleometrosis in the fire ant, Solenopsis invicta" (PDF). Мінез-құлық экологиясы және социобиология. 12 (2): 103–113. дои:10.1007/BF00343200. S2CID 3017347.
- ^ Trager 1988, б. 217.
- ^ а б Trager 1988, б. 218.
- ^ Manfredini, F.; Riba-Grognuz, O.; Wurm, Y.; Keller, L.; Shoemaker, D.; Grozinger, C.M.; Tsutsui, N.D. (2013). "Sociogenomics of cooperation and conflict during colony founding in the fire ant Solenopsis invicta". PLOS генетикасы. 9 (8): e1003633. дои:10.1371/journal.pgen.1003633. PMC 3738511. PMID 23950725.
- ^ Bernasconi, G.; Keller, L. (1996). "Reproductive conflicts in cooperative associations of fire ant queens (Solenopsis invicta)" (PDF). Корольдік қоғамның еңбектері B: Биологиялық ғылымдар. 263 (1369): 509–513. Бибкод:1996RSPSB.263..509B. дои:10.1098/rspb.1996.0077. S2CID 84276777.
- ^ Taber 2000, б. 42.
- ^ Doebeli, Mi.; Abouheif, E. (2016). "Modeling evolutionary transitions in social insects". eLife. 5: e12721. дои:10.7554/eLife.12721. PMC 4744194. PMID 26780668.
- ^ Hölldobler & Wilson 1990, б. 157.
- ^ Hölldobler & Wilson 1990, б. 168.
- ^ Markin, G. P.; Dillier, J. H.; Collins, H. L. (1973). "Growth and development of colonies of the red imported fire ant, Solenopsis invicta". Америка энтомологиялық қоғамының жылнамалары. 66 (4): 803–808. дои:10.1093/aesa/66.4.803.
- ^ а б Hölldobler & Wilson 1990, б. 163.
- ^ а б Taber 2000, б. 32.
- ^ Schmid-Hempel 1998, б. 10.
- ^ Schmid-Hempel 1998, б. 16.
- ^ Porter, S.D. (1988). "Impact of temperature on colony growth and developmental rates of the ant, Solenopsis invicta". Journal of Insect Physiology. 34 (12): 1127–1133. дои:10.1016/0022-1910(88)90215-6.
- ^ Helms, K.R.; Vinson, S.B. (2008). "Plant resources and colony growth in an invasive ant: the importance of honeydew-producing Hemiptera in carbohydrate transfer across trophic levels". Environmental Entomology. 37 (2): 487–493. дои:10.1093/ee/37.2.487. PMID 18419921.
- ^ Ross, K.G.; Fletcher, D.J.C. (1986). "Diploid male production — a significant colony mortality factor in the fire ant Solenopsis invicta (Hymenoptera: Formicidae) ». Мінез-құлық экологиясы және социобиология. 19 (4): 283–291. дои:10.1007/BF00300643. S2CID 25932018.
- ^ Ванг, Л .; Сю Ю .; Di, Z.; Roehner, B.M. (2013). "How does group interaction and its severance affect life expectancy?" (PDF). Корнелл университетінің кітапханасы: 1–28. arXiv:1304.2935. Бибкод:2013arXiv1304.2935W. S2CID 14848627.
- ^ Calabi, P.; Porter, S.D. (1989). "Worker longevity in the fire ant Solenopsis invicta: Ergonomic considerations of correlations between temperature, size and metabolic rates". Journal of Insect Physiology. 35 (8): 643–649. дои:10.1016/0022-1910(89)90127-3.
- ^ а б Hölldobler & Wilson 1990, б. 170.
- ^ Davies, N.B.; Krebs, J.R.; West, S.A. (2012). An Introduction to Behavioural Wcology (4-ші басылым). Чичестер: Уили-Блэквелл. б. 385. ISBN 978-1-4051-1416-5.
- ^ Passera, L.; Aron, S.; Vargo, E.L.; Keller, L. (2001). "Queen control of sex Ratio in fire ants" (PDF). Ғылым. 293 (5533): 1308–1310. Бибкод:2001Sci...293.1308P. дои:10.1126/science.1062076. PMID 11509728. S2CID 7483158.
- ^ Vargo, E.L.; Fletcher, D.J.C. (1986). "Evidence of pheromonal queen control over the production of male and female sexuals in the fire ant, Solenopsis invicta". Салыстырмалы физиология журналы А. 159 (6): 741–749. дои:10.1007/BF00603727. ISSN 1432-1351. S2CID 1165247.
- ^ а б Tschinkel 2006, б. 95.
- ^ Hölldobler & Wilson 1990, б. 187.
- ^ Goodisman, Michael A. D.; Ross, K.G. (1998). "A test of queen recruitment models using nuclear and mitochondrial markers in the fire ant Solenopsis invicta" (PDF). Эволюция. 52 (5): 1416–1422. дои:10.2307/2411311. JSTOR 2411311. PMID 28565383. Архивтелген түпнұсқа (PDF) 5 мамыр 2015 ж. Алынған 4 мамыр 2015.
- ^ Morel, L.; Meer, R.K.V.; Lofgren, C.S. (1990). "Comparison of nestmate recognition between monogyne and polygyne populations of Solenopsis invicta (Hymenoptera: Formicidae) ». Америка энтомологиялық қоғамының жылнамалары. 83 (3): 642–647. дои:10.1093/aesa/83.3.642.
- ^ Schmid-Hempel 1998, б. 191.
- ^ Trager 1988, б. 204.
- ^ Fletcher, D.J.C.; Блум, М.С .; Whitt, T.V.; Temple, N. (1980). "Monogyny and polygyny in the fire ant, Solenopsis invicta". Америка энтомологиялық қоғамының жылнамалары. 73 (6): 658–661. дои:10.1093/aesa/73.6.658.
- ^ Greenberg, L.; Fletcher, D.J.C.; Vinson, S.B. (1985). "Differences in worker size and mound distribution in monogynous and polygynous colonies of the fire ant Solenopsis invicta Buren". Канзас энтомологиялық қоғамының журналы. 58 (1): 9–18. JSTOR 25084596.
- ^ Harrison, R.G. (1980). "Dispersal polymorphisms in insects". Экология мен систематиканың жылдық шолуы. 11 (1): 95–118. дои:10.1146/annurev.es.11.110180.000523. JSTOR 2096904.
- ^ DeHeer, C.J.; Goodisman, M.A.D.; Ross, K.G. (1999). "Queen dispersal strategies in the multiple‐queen form of the fire ant Solenopsis invicta" (PDF). Американдық натуралист. 153 (6): 660–675. дои:10.1086/303205. ISSN 0003-0147. JSTOR 303205. Архивтелген түпнұсқа (PDF) 2015 жылғы 24 қыркүйекте. Алынған 4 мамыр 2015.
- ^ Keller, L. (1995). "Social life: the paradox of multiple-queen colonies". Экология мен эволюция тенденциялары. 10 (9): 355–360. дои:10.1016/S0169-5347(00)89133-8. PMID 21237068.
- ^ а б Ross, Kenneth G. (1993). "The breeding system of the fire ant Solenopsis invicta: Effects on colony genetic structure". Американдық натуралист. 141 (4): 554–76. дои:10.1086/285491. JSTOR 2462750. PMID 19425999.
- ^ Goodisman, M.A.D.; Sankovich, K.A.; Kovacs, J.L. (2006). "Genetic and morphological variation over space and time in the invasive fire ant Solenopsis invicta" (PDF). Biological Invasions. 9 (5): 571–584. дои:10.1007/s10530-006-9059-8. S2CID 25341015. Архивтелген түпнұсқа (PDF) 2015 жылғы 24 қыркүйекте. Алынған 4 мамыр 2015.
- ^ Adams, E.S.; Balas, M.T. (1999). "Worker discrimination among queens in newly founded colonies of the fire ant Solenopsis invicta". Мінез-құлық экологиясы және социобиология. 45 (5): 330–338. дои:10.1007/s002650050568. S2CID 23876496.
- ^ Chirino, M.G.; Gilbert, L.E.; Folgarait, P.J. (2012). "Behavioral discrimination between monogyne and polygyne red fire ants (Hymenoptera: Formicidae) in their native range". Америка энтомологиялық қоғамының жылнамалары. 105 (5): 740–745. дои:10.1603/AN11073. S2CID 86326110.
- ^ Goodisman, M.; Ross, K.G. (1997). "Relationship of queen number and queen relatedness in multiple-queen colonies of the fire ant Solenopsis invicta". Экологиялық энтомология. 22 (2): 150–157. дои:10.1046/j.1365-2311.1997.t01-1-00052.x.
- ^ Adams, E.S.; Tschinkel, W.R. (2001). "Mechanisms of population regulation in the fire ant Solenopsis invicta: an experimental study". Жануарлар экологиясының журналы. 70 (3): 355–369. дои:10.1046/j.1365-2656.2001.00501.x. S2CID 18657087.
- ^ Pracana, R.; Priyam, A.; Levantis, I.; Nichols, R.A.; Wurm, Y. (2017). "The fire ant social chromosome supergene variant Sb shows low diversity but high divergence from SB". Молекулалық экология. 26 (11): 2864–2879. дои:10.1111/mec.14054. PMC 5485014. PMID 28220980.
- ^ Ванг Дж .; Wurm, Y.; Nipitwattanaphon, M.; Riba-Grognuz, O.; Huang, Y.-C.; Shoemaker, D.; Keller, L. (2013). "A Y-like social chromosome causes alternative colony organization in fire ants" (PDF). Табиғат. 493 (7434): 664–668. Бибкод:2013 ж. 499..664W. дои:10.1038 / табиғат11832. PMID 23334415. S2CID 4359286.
- ^ Росс, К.Г .; Кригер, МДж .; Етікші, Д.Д. (2003). «От құмырсқаларындағы негізгі әлеуметтік полиморфизмнің альтернативті генетикалық негіздері». Генетика. 165 (4): 1853–1867. PMC 1462884. PMID 14704171.
- ^ «Қызыл импортталған өрт құмырсқасының әсерінен Австралияның табиғи фаунасы мен флорасының биоалуантүрлілігінің төмендеуі, Solenopsis invicta (от құмырсқасы) «. Қоршаған орта және энергетика бөлімі. Австралия үкіметі. Архивтелген түпнұсқа 2016 жылғы 22 қыркүйекте. Алынған 10 тамыз 2016.
- ^ а б в г. Войцик, Д .; Аллен, К.Р .; Бреннер, Р.Ж .; Форис, Э.А .; Джувеназ, Д.П .; Луц, Р.С. (2001). «Қызыл импортталған өрт құмырсқалары: биоәртүрлілікке әсері» (PDF). Американдық энтомолог. 47 (1): 16–23. дои:10.1093 / ae / 47.1.16. Архивтелген түпнұсқа (PDF) 16 желтоқсан 2010 ж.
- ^ Портер, С.Д .; Савиньяно, Д.А. (1990). «Полигинді отты құмырсқалардың шабуылы жергілікті құмырсқаларды жойып, буынаяқтылар қауымдастығын бұзады». Экология. 71 (6): 2095–2106. дои:10.2307/1938623. JSTOR 1938623. S2CID 56244004.
- ^ Стокер, Р.Л .; Грант, В.Е .; Брэдли Винсон, С. (1995). «Solenopsis invicta (Hymenoptera: Formicidae) Техастың орталық бөлігіндегі каррионның омыртқасыз ыдыратушыларына әсері ». Экологиялық энтомология. 24 (4): 817–822. дои:10.1093 / ee / 24.4.817.
- ^ Рикс, Б.Л .; Винсон, Б.С. (1970). «Белгілі бір жәндіктер мен әртүрлі суда еритін қосылыстардың импортталған өрт құмырсқасының екі сортына берілу қабілеті». Экономикалық энтомология журналы. 63 (1): 145–148. дои:10.1093 / jee / 63.1.145.
- ^ Кук, Дж.Л. (2003). «Қызыл импортталған өрт құмырсқасы әсер еткен аймақта биоәртүрлілікті сақтау, Solenopsis invicta (Hymenoptera: Formicidae) ». Биоалуантүрлілік және сақтау. 12 (2): 187–195. дои:10.1023 / A: 1021986927560. S2CID 28425331.
- ^ Массер, М.П .; Грант, В.Е. (1986). «Техастың шығыс-орталық бөлігіндегі ұсақ сүтқоректілердің отқа құмырсқалардан болатын өлім-жітім». Оңтүстік-Батыс натуралисті. 31 (4): 540–542. дои:10.2307/3671712. JSTOR 3671712.
- ^ Фликингер, Э.Л. (1989). «Тірі қалған жабайы мақта егеуқұйрықтарында қызыл импортталған өрт құмырсқалары арқылы жыртқыштықты бақылау». Texas Journal of Science. 41 (2): 223–224.
- ^ Хилл, Э.П. (1970). «Ұя салып жатқан мақта бүршіктеріндегі импортталған өрт құмырсқаларының жыртқыштықтарын бақылау». Жыл сайынғы конференция материалдары, аңшылық және балық комиссарларының оңтүстік-шығыс ассоциациясы. 23: 171–181.
- ^ Таули, Дж .; Лангкилде, Т. (2016). «Инвазиялық өрт құмырсқасы (Solenopsis invicta) шығыс қоршау кесірткесінің жыртқыштығы (Sceloporus undulatus) жұмыртқа ». Герпетология журналы. 50 (2): 284–288. дои:10.1670/15-017. S2CID 85855854.
- ^ Маунт, Р.Х .; Траут, С.Е .; Мейсон, В.Х. (1981). «Қызыл импортталған өрт құмырсқасының жыртқыштығы, Solenopsis invicta (Hymenoptera: Formicidae), кесіртке жұмыртқаларында Cnemidophorus sexlineatus (Squamata: Teiidae) ». Алабама ғылым академиясының журналы. 52: 66–70.
- ^ Бартлетт, Д. (1997). «Пейнс Прериясы туралы 40 жылдық ойлар». Бауырымен жорғалаушылар. 5 (7): 68, 70–73.
- ^ Лангкилде, Т. (2009). «Инвазиялық от құмырсқалары жергілікті кесірткелердің мінез-құлқы мен морфологиясын өзгертеді». Экология. 90 (1): 208–217. дои:10.1890/08-0355.1. PMID 19294926. S2CID 670780.
- ^ Брайнер, Дж. (26 қаңтар 2009). «Кесірткелердің биі өлімге толы құмырсқадан аулақ болады». LiveScience. Алынған 20 тамыз 2016.
- ^ Босатылды, P.S .; Нейтман, К. (1988). «Жойылу қаупі төніп тұрған Хьюстон құрбақасындағы жыртқыштық туралы ескертулер, Bufo houstonensis". Texas Journal of Science. 40: 454–456.
- ^ Аллен, К.Р .; Райс, К.Г .; Войцик, Д.П .; Percival, H.F. (1997). «Қызыл түсті импортталған отты құмырсқаны инвеномизациялаудың жаңа туылған американдық аллигаторларға әсері». Герпетология журналы. 31 (2): 318–321. дои:10.2307/1565408. JSTOR 1565408.
- ^ а б Capinera 2008, б. 1080.
- ^ Кастелланос, Адриан А .; Медерос, Мэттью C. Мен .; Хамер, Габриэль Л .; Морроу, Майкл Э .; Эубанкс, Мики Д .; Тил, Пит Д .; Хамер, Сара А .; Light, Jessica E. (2016). «Инвазиялық қызыл импортталған өрт құмырсқаларымен байланысты ұсақ сүтқоректілер мен қожайындардың кенесінің азаюы (Solenopsis invicta)". Биология хаттары. 12 (9): 20160463. дои:10.1098 / rsbl.2016.0463 ж. PMC 5046925. PMID 27651533.
- ^ Молони, С.Д .; Vanderwoude, C. (2003). «Австралияның шығысындағы қызыл импортталған өрт құмырсқаларының экологиялық әсері» (PDF). Ауылшаруашылық және қалалық энтомология журналы. 20 (3): 131–142.
- ^ Локли, ТК (1995). «Шетелден әкелінетін өрт құмырсқасының жыртқыштықтың ең аз террогия популяциясына әсері: жойылып бара жатқан түр». Оңтүстік-батыс энтомологы. 20: 517–519.
- ^ Сикес, П.Ж .; Арнольд, К.А. (1986). «Қызыл импортталған өрт құмырсқасы (Solenopsis invicta) жартас қарлығашындағы жыртқыштық (Хирундо пирронота) орталық Техас штатындағы ұялар ». Оңтүстік-батыс натуралисті. 31 (1): 105–106. дои:10.2307/3670967. JSTOR 3670967.
- ^ Кампомицци, А.Ж .; Моррисон, М .; Фаррелл, С.Л .; Уилкинс, Р..Н .; Дризес, Б.М .; Packard, JM (2009). «Қызыл импортталған өрт құмырсқалары ән құсының ұясының тіршілігін төмендетуі мүмкін». Кондор. 111 (3): 534–537. дои:10.1525 / cond.2009.090066. S2CID 84552544.
- ^ Морроу, М.Е .; Честер, Р.Е .; Лехен, С.Е .; Дризес, Б.М .; Toepfer, J.E. (2015). «Қызыл импортталған өрт құмырсқаларының Attwater-дың дала-тауық тұқымдарының тірі қалуына жанама әсері». Жабайы табиғатты басқару журналы. 79 (6): 898–906. дои:10.1002 / jwmg.915. PMC 4745021. PMID 26900176.
- ^ Йосеф, Р .; Лорер, Ф.Е. (1995). «Логгерхед ұрады, қызыл от құмырсқалары мен қызыл майшабақтар?» (PDF). Кондор. 97 (4): 1053–1056. дои:10.2307/1369544. JSTOR 1369544.
- ^ «Қоршаған орта: Отпен күресу». Уақыт. 2 қараша 1970. мұрағатталған түпнұсқа 2016 жылғы 22 қыркүйекте. Алынған 28 тамыз 2016.
- ^ Capinera 2008, б. 1771.
- ^ Кинг, Дж .; Цчинкель, В.Р. (2006). «Эксперименттік дәлелдеме, өрт сөндіру құмырасы енгізілген Solenopsis invicta, бұзылған тіршілік ету ортасында бірге пайда болатын құмырсқаларды бәсекеге жол бермейді ». Жануарлар экологиясының журналы. 75 (6): 1370–1378. дои:10.1111 / j.1365-2656.2006.01161.x. JSTOR 4125078. PMID 17032369. S2CID 14719552.
- ^ Гао, Ю .; Лу, Л-Х .; Ол, Y-R .; Ци, Дж-Дж .; Чжан, Дж. (2011). «Қызыл импортталған өрт құмырсқасы арасындағы кедергі бәсекелестігі (Solenopsis invicta Buren) және екі құмырсқа түрі (Hymenoptera: Formicidae) ». Acta Entomologica Sinica. 54 (5): 602–608.
- ^ Лебрун, Э.Г .; Эбботт, Дж .; Гилберт, Л.Е. (2013). «Импортталған ессіз құмырсқа импортталған өрт құмырсқасын ығыстырады, шабындық құмырсқалар мен буынаяқтылардың жиынтығын азайтады және біртектес етеді». Биологиялық инвазиялар. 15 (11): 2429–2442. дои:10.1007 / s10530-013-0463-6. S2CID 15260861.
- ^ Витон, Т .; Lucky, A. (2014). «Тротуар құмырсқасы - Tetramorium Caespitum". UF / IFAS таңдаулы жаратылыстар. Флорида университеті. Алынған 4 сәуір 2016.
- ^ King, TG .; Филлипс, СА (1992). «Қызыл колониялардың әкелінетін қызыл колонияларды тротуар құмырсқасы арқылы жою». Энтомологиялық жаңалықтар. 103 (3): 72–77. ISSN 0013-872X.
- ^ Чен, Ю.К .; Кафле, Л .; Shih, CJ (2011). «Арасындағы түраралық бәсекелестік Solenopsis invicta және екі құмырсқа түрі, Феидол фервендері және Monomorium chinense". Экономикалық энтомология журналы. 104 (2): 614–621. дои:10.1603 / ec10240. PMID 21510213. S2CID 44631810.
- ^ Калькатерра, Л.А .; Ливоре, Дж.П .; Делгадо, А .; Брайано, Дж.А. (2008). «Қызыл импортталған өрт құмырсқасының экологиялық үстемдігі, Solenopsis invicta, өзінің диапазонында ». Oecologia. 156 (2): 411–421. Бибкод:2008Oecol.156..411C. дои:10.1007 / s00442-008-0997-ж. PMID 18305962. S2CID 21842183.[тұрақты өлі сілтеме ]
- ^ Лебрун, Э.Г .; Тиллберг, C.V .; Суарес, А.В .; Фолгарайт, П.Ж .; Смит, Кр .; Холуэй, Д.А. (2007). «От құмырсқалары мен аргентиналық құмырсқалардың өздерінің таралу аймағында бәсекелестігін эксперименттік зерттеу» (PDF). Экология. 88 (1): 63–75. дои:10.1890 / 0012-9658 (2007) 88 [63: AESOCB] 2.0.CO; 2. PMID 17489455.
- ^ Чжоу, А .; Лян, Г .; Лу, Ю .; Ценг, Л .; Xu, Y. (2014). «Қызыл импортталған өрт құмырсқасы арасындағы түраралық бәсекелестік, Solenopsis invicta Бүрен мен елес құмырсқа, Тапинома меланоцефалия Инвазивті мелебогпен жасалған бал шірі ресурстарына арналған Fabricius, Фенакокк соленопсисі Тинсейли »тақырыбында өтті. Буынаяқтылар-өсімдіктердің өзара әрекеттесуі. 8 (5): 469–474. дои:10.1007 / s11829-014-9324-1. S2CID 18102670.
- ^ Ву, Б .; Ванг, Л .; Лян, Г .; Лу, Ю .; Zeng, L. (2014). «Арасындағы азық-түлік бәсекелестігінің механизмі Solenopsis invicta Бүрен және Тапинома меланоцефалия Фабрициус «. Әлеуметтану. 61 (3): 265–273. дои:10.13102 / социобиология.v61i3.265-273. Архивтелген түпнұсқа 2016 жылғы 22 қыркүйекте.
- ^ Кассилл, Д.Л .; Во, К .; Беккер, Б. (2008). «Өрт сөндіретін жас жұмысшылар өлімді сезінеді және агрессивті көршілерінен аман қалады». Naturwissenschaften. 95 (7): 617–624. Бибкод:2008NW ..... 95..617C. дои:10.1007 / s00114-008-0362-3. PMID 18392601. S2CID 2942824.
- ^ Кабашима, Дж.Н .; Гринберг, Л .; Rust, M.K .; Пейн, ТД (2007). «Арасындағы агрессивті өзара әрекеттесу Solenopsis invicta және Линепитема кішіпейілді (Hymenoptera: Formicidae) зертханалық жағдайда « (PDF). Экономикалық энтомология журналы. 100 (1): 148–154. дои:10.1603 / 0022-0493 (2007) 100 [148: AIBSIA] 2.0.CO; 2. PMID 17370822.
- ^ Capinera 2008, б. 290.
- ^ а б Чжоу, А .; Лу, Ю .; Ценг, Л .; Сю Ю .; Лианг, Г. (2012). «Мутуализм екі жат түрдің шабуылына себеп бола ма? Solenopsis invicta және Фенакокк соленопсисі". PLOS ONE. 7 (7): e41856. Бибкод:2012PLoSO ... 741856Z. дои:10.1371 / journal.pone.0041856. PMC 3402455. PMID 22911859.
- ^ Каплан, Мен .; Eubanks, MD (2002). «Мақта апидінің бұзылуы (Homoptera: Aphididae) - қызыл импортталған өрт құмырсқаларының табиғи жау динамикасы (Hymenoptera: Formicidae)». Экологиялық энтомология. 31 (6): 1175–1183. дои:10.1603 / 0046-225X-31.6.1175. S2CID 34957731.
- ^ Копплер, Л.Б .; Мерфи, Дж. Ф .; Eubanks, MD (2007). «Қызыл импортталған өрт құмырсқалары (Hymenoptera: Formicidae) қызанақтағы тлидің көптігін арттырады». Флорида энтомологы. 90 (3): 419–425. дои:10.1653 / 0015-4040 (2007) 90 [419: RIFAHF] 2.0.CO; 2.
- ^ Диас, Р .; Кнутсон, А .; Бернал, Дж. (2004). «Қызыл импортталған өрт құмырсқасының мақта тлиінің популяциясының тығыздығына және құрт пен қызылша армия құрттарының жұмыртқаларына әсері». Экономикалық энтомология журналы. 97 (2): 222–229. дои:10.1603/0022-0493-97.2.222. PMID 15154439. S2CID 198130004.
- ^ Теддерлер, В.Л .; Рейли, СС .; Wood, B.W .; Моррисон, Р.К .; Лофгрен, СС (1990). «Мінез-құлық Solenopsis invicta (Hymenoptera: Formicidae) пекан бақтарында ». Экологиялық энтомология. 19 (1): 44–53. дои:10.1093 / ee / 19.1.44.
- ^ Табер, Стивен (тамыз 2000). От құмырсқалары (1-ші басылым). Texas A & M University Press. б. 308. ISBN 0890969450
- ^ Толығырақ, Даниэль Р .; Кольмейер, Р. Е .; Хоффман, Дональд Р. (наурыз 2008). «Нәресте ішіндегі отқа арналған құмырсқалардың шағуына өлімге әкелетін анафилаксия». Американдық сот медицинасы және патология журналы. 29 (1): 62–63. дои:10.1097 / PAF.0b013e3181651b53. PMID 19749619.
- ^ Фицджералд, Кевин Т .; Тасқын, Арын А. (қараша 2006). «Hymenoptera Stings». Кішкентай жануарлар практикасындағы клиникалық әдістер. 21 (4): 194–204. дои:10.1053 / j.ctsap.2006.10.002. PMID 17265905.
- ^ Роудс, Роберт Б .; Стаффорд, Честер Т .; Джеймс, Фрэнк К. (тамыз 1989). «Импортталған өрт құмырсқасының шағуына өлімге әкелетін анафилактикалық реакцияларды зерттеу». Аллергия және клиникалық иммунология журналы. 84 (2): 159–162. дои:10.1016/0091-6749(89)90319-9. PMID 2760357.
- ^ Замит-Миранда, Даниэль; Фокс, Эдуардо Г. П .; Монтейро, Ана Паула; Гама, Диого; Публан, Луис Е .; де Араудо, Алмаир Феррейра; Арауо, Мария Ф. С .; Ателла, Джорджия С .; Мачадо, Эднилдо А .; Диас, Бруно Л. (26 қыркүйек 2018). «Өрт құмырсқасының уы ақуыздарының көмегімен аллергиялық жауап». Ғылыми баяндамалар. 8 (1): 14427. Бибкод:2018 Натрия ... 814427Z. дои:10.1038 / s41598-018-32327-з. PMC 6158280. PMID 30258210.
- ^ Fox E.G.P. (2016 ж.) От құмырсқаларының уы токсиндері. In: Gopalakrishnakone P., Calvete J. (eds) Venom Genomics and Proteomics. Токсинология. Спрингер, Дордрехт
- ^ дос Сантос Пинто, Хосе Р. А .; Фокс, Эдуардо Г. П .; Саидемберг, Даниэль М .; Сантос, Люсилен Д .; да Силва Менегассо, Аналли Р .; Коста-Мансо, Элиуда; Мачадо, Эднилдо А .; Буэно, Одаир С .; Пальма, Марио С. (22 тамыз 2012). «Оттың протеомиялық көрінісі отты құмырсқа Буреннен». Протеомды зерттеу журналы. 11 (9): 4643–4653. дои:10.1021 / pr300451g. PMID 22881118.
- ^ Баер, Х .; Лю, Т.-Ы .; Андерсон, МС .; Блум, М .; Шмид, В.Х .; Джеймс, Ф.Ж. (1979). «Өрт құмырсқасы уының ақуыздық компоненттері (Solenopsis invicta)". Токсикон. 17 (4): 397–405. дои:10.1016/0041-0101(79)90267-8. PMID 494321.
- ^ Хофман, DR; Көгершін, DE; Джейкобсон, RS (қараша 1988). «Hymenoptera уындағы аллергендер. ХХ. Импортталған өрт құмырсқасынан (Solenopsis invicta) төрт аллергенді оқшаулау». Аллергия және клиникалық иммунология журналы. 82 (5 Pt 1): 818-27. дои:10.1016 / 0091-6749 (88) 90084-X. PMID 3192865.
- ^ McDonald, M. (2006). «Аяқ астындағы қызылдар». Жаңа ғалым. 189 (2538): 50. мұрағатталған түпнұсқа 2016 жылғы 25 сәуірде.
- ^ а б в г. «Қызыл импортталған өрт құмырсқасы: Батыс Австралияда зиянкестер деп жарияланды». Ауыл шаруашылығы және тамақ бөлімі. Батыс Австралия үкіметі. 2014. мұрағатталған түпнұсқа 2016 жылғы 22 қыркүйекте. Алынған 21 тамыз 2016.
- ^ Пиментель, Д .; Лах, Л .; Зунига, Р .; Моррисон, Д. (2000). «АҚШ-тағы жергілікті емес түрлердің экологиялық және экономикалық шығындары». BioScience. 50 (1): 53–65. дои:10.1641 / 0006-3568 (2000) 050 [0053: EAECON] 2.3.CO; 2.
- ^ Dye, J. (8 желтоқсан 2014). «Сиднейдегі қызыл өрт құмырсқасының шығуы экономикаға миллиардтаған шығын әкелуі мүмкін». Sydney Morning Herald. Алынған 8 желтоқсан 2014.
- ^ а б в «От құмырсқалары». Виктория. Ауыл шаруашылығы. Виктория үкіметі. 2004. мұрағатталған түпнұсқа 2016 жылғы 22 қыркүйекте. Алынған 20 тамыз 2016.
- ^ Смит, МР (1965). Америка Құрама Штаттарының үйге жұққан құмырсқалары: оларды тану, биология және экономикалық маңыздылығы (PDF). Вашингтон, Колумбия округі: Америка Құрама Штаттарының Ауыл шаруашылығы министрлігі. б. 43. OCLC 6078460.
- ^ Стайлз, Дж. Х .; Джонс, РХ (1998). «Қызыл импортталған өрт құмырсқасының таралуы, формасы Solenopsis invicta, электр және электр желілерінің тіршілік ету орталарында ». Ландшафттық экология. 13 (6): 335–346. дои:10.1023 / A: 1008073813734. S2CID 21131952.
- ^ «АҚШ отты құмырсқалардың жауларын импорттайды. CNN. 30 қараша 1999 ж. Алынған 1 қараша 2016.
- ^ Вандер Меер, Р.К. Вандер; Словик, Т. Дж .; Торвилсон, Х. Г. (2002). «Электрмен қоздырылған қызыл импортталған отты құмырсқалар шығаратын жартылай химикаттар, Solenopsis invicta". Химиялық экология журналы. 28 (12): 2585–2600. дои:10.1023 / A: 1021448522147. PMID 12564802. S2CID 4705569.
- ^ Маккей, В.П .; Винсон, С.Б .; Ирвинг, Дж .; Маджди, С .; Мессер, C. (1992). «Электр өрістерінің қызыл импортталған өрт құмырсқасына әсері (Hymenoptera: Formicidae)». Экологиялық энтомология. 21 (4): 866–870. дои:10.1093 / ee / 21.4.866.
- ^ Словик, Т.Ж .; Жасыл, Б.Л .; Торвилсон, Х.Г. (1997). «Қызыл импортталған өрт құмырсқасында магниттілікті анықтау (Solenopsis invicta) магниттік-резонанстық томографияны қолдану ». Биоэлектромагнитика. 18 (5): 396–399. дои:10.1002 / (SICI) 1521-186X (1997) 18: 5 <396 :: AID-BEM7> 3.0.CO; 2-Y. PMID 9209721.
- ^ Андерсон, Дж.Б .; Вандер Меер, Р.К. (1993). «Өрт құмырсқасындағы магниттік бағдар, Solenopsis invicta". Naturwissenschaften. 80 (12): 568–570. Бибкод:1993NW ..... 80..568A. дои:10.1007 / BF01149274. S2CID 41502040.
- ^ а б Джеттер, К.М .; Гамильтон, Дж .; Клотц, Дж. (2002). «Жою шығындары есептелді: Қызыл импортталған өрт құмырсқалары ауыл шаруашылығына, жануарлар дүниесіне және үйлерге қауіп төндіреді» (PDF). Калифорния ауыл шаруашылығы. 56 (1): 26–34. дои:10.3733 / ca.v056n01p26.
- ^ «Отқа құмырсқалардың әсері - неге олар проблема болып табылады». Ауыл шаруашылығы және балық шаруашылығы бөлімі. Квинсленд үкіметі. 2016. мұрағатталған түпнұсқа 2016 жылғы 22 қыркүйекте. Алынған 20 тамыз 2016.
- ^ Лофгрен, СС .; Адамс, С.Т. (1981). «Қызыл импортталған өрт құмырсқасы егіс алқаптарындағы сояның өнімділігі төмендеді, Solenopsis invicta". Флорида энтомологы. 64 (1): 199–202. дои:10.2307/3494619. JSTOR 3494619.
- ^ Банктер, В.А .; Адамс, К.Т .; Лофгрен, СС .; Войцик, Д.П. (1990). «Құрама Штаттардың оңтүстігінде соя алқабына импортталған өрт құмырсқасы». Флорида энтомологы. 73 (3): 503–504. дои:10.2307/3495467. JSTOR 3495467.
- ^ Адамс, К.Т .; Пламли, Дж .; Лофгрен, СС .; Банктер, АҚШ (1976). «Қызыл импортталған өрт құмырсқасының экономикалық маңызы, Solenopsis invicta Бөрен. I. Сояның жинауына әсерін алдын-ала тергеу. Джорджия энтомологиялық қоғамының журналы. 11: 165–169.
- ^ Адамс, К.Т .; Пламли, Дж .; Банктер, В.А .; Лофгрен, С.С. (1977). «Қызыл импортталған өрт құмырсқасының әсері, Solenopsis invicta Бурен (Hymenoptera: Formicidae) Солтүстік Каролинадағы соя дақылын жинау туралы ». Элиша Митчелл қоғамының журналы. 93: 150–153.
- ^ Апперсон, СС .; Адамс, С.Т. (1983). «Қызыл импортталған өрт құмырсқасының медициналық және ауылшаруашылық маңызы». Флорида энтомологы. 66 (1): 121–126. дои:10.2307/3494558. JSTOR 3494558.
- ^ Capinera 2008, б. 903.
- ^ Банктер, В.А .; Адамс, К.Т .; Лофгрен, C.S. (1991). «Қызыл цитрус ағаштарына қызыл әкелінген өрт құмырсқасының зақымдануы (Hymenoptera: Formicidae)». Экономикалық энтомология журналы. 84 (1): 241–246. дои:10.1093 / jee / 84.1.241.
- ^ Capinera 2008, б. 1084.
- ^ Стерлинг, В.Л. (1978). «Қызыл импортталған өрт құмырсқасы арқылы қоңыздар тұқымын биологиялық жолмен басу». Экологиялық энтомология. 7 (4): 564–568. дои:10.1093 / ee / 7.4.564.
- ^ Филлман, Д.А .; Стерлинг, В.Л. (1983). «Қызыл импортталған өрт құмырсқасының өлтіру күші [Hym .: Formicidae]: қарақұйрықтың негізгі жыртқышы [Col. Curculionidae]». Энтомофага. 28 (4): 339–344. дои:10.1007 / BF02372186. S2CID 38550501.
- ^ Стерлинг, В.Л .; Дин, Д.А .; Филлман, Д.А .; Джонс, Д. (1984). «Сыбыргүлділердің табиғи биологиялық бақылауы [Col. Curculionidae]». Энтомофага. 29 (1): 1–9. дои:10.1007 / BF02372203. S2CID 33045477.
- ^ Негм, А.А .; Хенсли, С.Д. (1967). «Буынаяқтылардың жыртқыштарының қант қамышының қопсытқышымен келтірілген дақылдардың зақымдануына қатынасы». Экономикалық энтомология журналы. 60 (6): 1503–1506. дои:10.1093 / jee / 60.6.1503.
- ^ Рейган, Т.Е .; Кобурн, Г .; Хенсли, С.Д. (1972). «Мирекстің Луизиана қант қамысы өрісінің артроподты фаунасына әсері». Экологиялық энтомология. 1 (5): 588–591. дои:10.1093 / ee / 1.5.588.
- ^ Capinera 2008, б. 4173.
- ^ Лемке, Л.А .; Kissam, JB (1988). «Про-дронмен өңделген мал жайылымында қызыл импортталған отты құмырсқалардың (Hymenoptera: Formicidae) мүйіз шыбындарға әсері» (Diptera: Muscidae). Экономикалық энтомология журналы. 81 (3): 855–858. дои:10.1093 / jee / 81.3.855. PMID 3403766.
- ^ Ли, Дж. Джонсон, С.Ж .; Райт, В.Л. (1990). «Луизианадағы соя егістігінде барқыт құртының (Lepidoptera: Noctuidae) қуыршақтарын тірі қалудың сандық талдауы». Экологиялық энтомология. 19 (4): 978–986. дои:10.1093 / ee / 19.4.978.
- ^ Моррилл, В.Л. (1977). «Жылыжайда қызыл импортталған өрт құмырсқасы». Экологиялық энтомология. 6 (3): 416–418. дои:10.1093 / ee / 6.3.416.
- ^ а б в Eubanks, MD (2001). «Қызыл импортталған өрт құмырсқаларының егістік дақылдарындағы биологиялық бақылауға тікелей және жанама әсерін бағалау». Биологиялық бақылау. 21 (1): 35–43. дои:10.1006 / bcon.2001.0923 ж. S2CID 86682078.
- ^ Уилсон, Н.Л .; Olive, AD (1969). «Луизиана штатының оңтүстік-шығысында жайылымдық және қарағайлы орман алқаптарында әкелінген от құмырсқасының тамақтану әдеттері». Экономикалық энтомология журналы. 62 (6): 1268–1271. дои:10.1093 / jee / 62.6.1268.
- ^ Ху, Г.Я .; Фрэнк, Дж. (1996). «Қызыл импортталған өрт құмырсқасының (Hymenoptera: Formicidae) Флоридадағы тезек мекендейтін буынаяқтыларға әсері». Экологиялық энтомология. 25 (6): 1290–1296. дои:10.1093 / ee / 25.6.1290.
- ^ Лопес, ДжД (1982). «Қыстайтын халықтың пайда болу заңдылығы Нигрицептердің кардиохилдері орталық Техаста ». Экологиялық энтомология. 11 (4): 838–842. дои:10.1093 / ee / 11.4.838.
- ^ Elliot, D. (1 шілде 2013). ""Crazy құмырсқалар «оңтүстік-шығыс штаттарда». CBS жаңалықтары. Алынған 22 қараша 2016.
- ^ Лофгрен, СС .; Банктер, В.А .; Глэнси, Б.М. (1975). «Импортталған өрт құмырсқаларының биологиясы және бақылау». Энтомологияның жылдық шолуы. 20 (1): 1–30. дои:10.1146 / annurev.en.20.010175.000245. PMID 1090234.
- ^ Бухс 2005 ж, 1-2 беттер.
- ^ Уильямс, Д.Ф .; deShazo, RD (2004). «От құмырсқаларын биологиялық бақылау: жаңа техниканы жаңарту». Аллергия, астма және иммунология жылнамалары. 93 (1): 15–22. дои:10.1016 / S1081-1206 (10) 61442-1. PMID 15281467.
- ^ Mottern, J.L; Хайнц, К.М; Ode, PJ (2004). «Өрттен жасалған құмырсқаларды фридті шыбындармен биологиялық бақылауды бағалау: бәсекелік өзара әрекеттесуге әсер ету». Биологиялық бақылау. 30 (3): 566–583. дои:10.1016 / j.biocontrol.2004.02.006.
- ^ Дризес, Б.М .; Пакетт, Р. «Қызыл импортталған отты құмырсқаға қарсы биологиялық бақылау құралдары» (PDF). Техас импортталған өрт қауырсындарын зерттеу және басқару жобасы. Texas A&M University. Алынған 21 тамыз 2016.
- ^ Бурен, В.Ф .; Аллен, Дж .; Уильямс, Р.Н. (1978). «Импортталған өрт құмырсқаларын зиянкестермен басқарудың тәсілдері». Американың энтомологиялық қоғамының хабаршысы. 24 (4): 418–421. дои:10.1093 / besa / 24.4.418.
- ^ а б в «Зиянкестермен күресудің интеграцияланған нұсқаулығы: өрт сөндіргіштер». Ұлттық парк қызметі. Америка Құрама Штаттарының ішкі істер департаменті. 2010. мұрағатталған түпнұсқа 2016 жылғы 22 қыркүйекте. Алынған 21 тамыз 2016.
- ^ Боннефой, Х .; Кампен, Х .; Суини, К. (2008). Қалалық зиянкестердің денсаулық сақтаудың маңызы. Копенгаген, Дания: Дүниежүзілік денсаулық сақтау ұйымы, Еуропалық аймақтық бюро. б. 193. ISBN 978-92-890-7188-8.
- ^ Каликто, А.А .; Харрис, М.К .; Кнутсон, А .; Барр, Калифорния (2007). «Жергілікті құмырсқалардың жауаптары Solenopsis invicta Таратылған жемдерді қолдану арқылы редукцияны азайту ». Экологиялық энтомология. 36 (5): 1112–1123. дои:10.1603 / 0046-225X (2007) 36 [1112: NARTSI] 2.0.CO; 2. PMID 18284735.
- ^ Клотц, Дж. Х .; Вэйл, К.М .; Уилламс, Д.Ф. (1997). «Бор қышқылы-сахарозаның су жемінің уыттылығы Solenopsis invicta (Hymenoptera: Formicidae) ». Экономикалық энтомология журналы. 90 (2): 488–491. дои:10.1093 / jee / 90.2.488.
- ^ Бенсон, Э.П .; Зунголи, П.А .; Райли, М.Б. (2003). «Ластауыштардың жемді қабылдауға әсері Solenopsis invicta (Hymenoptera: Formicidae) ». Экономикалық энтомология журналы. 96 (1): 94–97. дои:10.1093 / jee / 96.1.94. PMID 12650350.
Библиография
- Buhs, JB (2005). От құмырсқалары: ХХ ғасырдағы Америкадағы табиғат, ғылым және мемлекеттік саясат. Чикаго, Иллинойс: Чикаго университеті баспасы. ISBN 978-0-226-07981-3.CS1 maint: ref = harv (сілтеме)
- Capinera, JL (2008). Энтомология энциклопедиясы (2-ші басылым). Дордрехт, Нидерланды: Шпрингер. ISBN 978-1-4020-6242-1.CS1 maint: ref = harv (сілтеме)
- Хёльдоблер, Б .; Уилсон, Э.О. (1990). Құмырсқалар. Кембридж, Массачусетс: Гарвард университетінің Белнап баспасы. ISBN 978-0-674-04075-5.CS1 maint: ref = harv (сілтеме)
- Schmid-Hempel, P. (1998). Әлеуметтік жәндіктердегі паразиттер. Принстон, Нью-Джерси: Принстон университетінің баспасы. ISBN 978-0-691-05924-2.CS1 maint: ref = harv (сілтеме)
- Табер, С.В. (2000). От құмырсқалары. College Station, Texas: Texas A&M University Press. ISBN 978-1-60344-711-9.CS1 maint: ref = harv (сілтеме)
- Trager, JC (1988). Мирмекологияның жетістіктері (1-ші басылым). Лейден, Нидерланды: Э.Дж. Брилл. ISBN 978-0-916846-38-1.CS1 maint: ref = harv (сілтеме)
- Цчинкель, В. (2006). От құмырсқалары. Кембридж, Массачусетс: Гарвард университетінің Белнап баспасы. ISBN 978-0-674-02207-2.CS1 maint: ref = harv (сілтеме)
Сыртқы сілтемелер
| Кітапхана қоры туралы Қызыл импортталған өрт құмырсқасы |
Веб-сайт
- Қызыл импортталған от құмырсқасы туралы кітаптар мен журналдар кезінде Биоалуантүрлілік мұралары кітапханасы
- Қызыл импортталған от құмырсқасы туралы кітаптар мен журналдар кезінде Интернет мұрағаты
- Ұлттық қызыл импортталған отты құмырсқаны жою бағдарламасы - Квинслендтің ауыл шаруашылығы және балық шаруашылығы департаменті
- Импортталған өрт құмырсқалары - Америка Құрама Штаттарының Ауыл шаруашылығы министрлігі
- Solenopsis invicta AntWiki-де - құмырсқаларды әлемге әкелу
- The S. invicta Геном жобасы - Hymenoptera Genome Database жобасы
- Түрлер профилі - қызыл импортталған өрт құмырсқасы (Solenopsis invicta), Ұлттық инвазиялық түрлер туралы ақпарат орталығы, Америка Құрама Штаттарының ұлттық ауылшаруашылық кітапханасы.
БАҚ
- Өртке қарсы құмырсқаларды паразиттейтін форид шыбындарының кадрлары - Техас Университеті Остин жаратылыстану ғылымдары колледжі шығарған BBC-мен түсірілген
- Қызыл от құмырсқалары - YouTube-те National Geographic жариялады
- Балқытылған алюминиймен отты құмырсқа колониясын құю YouTube-те
Жаңалықтар
- Керемет өзгермелі отты құмырсқа – Washington Post, 2011