Фитопланктон - Phytoplankton

| Серияның бір бөлігі |
| Планктон |
|---|
 |
Фитопланктон (/ˌfaɪтoʊˈблæŋктən/) болып табылады автотрофты компоненттері планктон қауымдастық және мұхит пен тұщы судың негізгі бөлігі экожүйелер. Атауы Грек сөздер φυτόν (фитон), мағынасы «өсімдік «, және πλαγκτός (планктоздар), «қаңғыбас» немесе «дрейфер» деген мағынаны білдіреді.[1]
Фитопланктон өзінің энергиясын алады фотосинтез, құрлықтағы ағаштар мен басқа өсімдіктер сияқты. Демек, фитопланктонның күн сәулесі болуы керек, сондықтан олар жақсы жарықтандырылған беткі қабаттарда өмір сүреді (эйфотикалық аймақ ) мұхиттар мен көлдер. Құрлықтағы өсімдіктермен салыстырғанда фитопланктон үлкен беткейге таралған, аз маусымдық өзгеріске ұшырайды және ағаштармен салыстырғанда жылдам айналым жылдамдығына ие (онжылдықтармен салыстырғанда күндер). Нәтижесінде фитопланктон ғаламдық ауқымда климаттың өзгеруіне тез жауап береді.
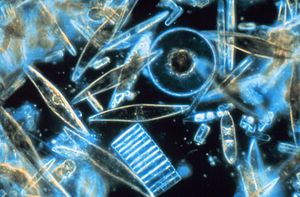
Фитопланктон теңіз және тұщы су қоректену торларының негізін құрайды және әлемдегі басты ойыншылар болып табылады көміртегі айналымы. Олардың үлесіне бүкіләлемдік фотосинтетикалық белсенділіктің жартысы және оттегі өндірісінің жартысына жуығы жатады, дегенмен бұл өсімдіктердің әлемдік биомассасының шамамен 1% құрайды. Фитопланктон өте алуан түрлі, олар фотосинтездейтін бактериялардан өсімдік тәрізді балдырларға дейін, броньмен қапталғанға дейін кокколитофорлар. Фитопланктонның маңызды топтарына мыналар жатады диатомдар, цианобактериялар және динофлагеллаттар дегенмен, көптеген басқа топтар ұсынылған.
Фитопланктондардың көпшілігі өте кішкентай, оларды жеке-жеке көру мүмкін емес көзсіз көз. Алайда, жеткілікті жоғары мөлшерде болған кезде, кейбір сорттардың болуы су бетіндегі түрлі-түсті дақтар ретінде байқалуы мүмкін хлорофилл олардың жасушаларында және аксессуар пигменттерінде (мысалы фикобилипротеидтер немесе ксантофилдер ) кейбір түрлерінде.
Түрлері
Фитопланктон болып табылады фотосинтездеу Жердегі барлық дерлік мұхиттар мен тұщы су айдындарының күн сәулесімен жарқыраған жоғарғы қабатын мекендейтін микроскопиялық биотикалық организмдер. Олар агенттер алғашқы өндіріс, құру органикалық қосылыстар бастап Көмір қышқыл газы суда еріген, суда тіршілік ететін процесс тамақтану торы.[2] Фитопланктон негізін құрайды теңіз тағамдары торы және олар Жердің шешуші ойыншылары болып табылады көміртегі айналымы.[3]

Фитопланктон фотосинтездейтін бактериялардан (цианобактериялардан) бастап өсімдік тәрізді диатомаларға дейін, броньмен қапталған кокколитофорларға дейін өте алуан түрлі.[4]


Экология

* Қызыл = диатомдар (кремний диоксиді қажет болатын үлкен фитопланктон)
* Сары = флагелаттар (басқа үлкен фитопланктон)
* Жасыл = прохлорококк (нитратты қолдана алмайтын кішкентай фитопланктон)
* Көгілдір = синехококк (басқа фитопланктон)
Мөлдірлігі көміртегі биомассасының концентрациясын көрсетеді. Атап айтқанда, мұхиттағы биологиялық әртүрлілікті сақтауда бұралулар мен жіптердің (мезоскальдік ерекшеліктер) рөлі маңызды болып көрінеді.[3][5]
Фитопланктон алады энергия арқылы процесс туралы фотосинтез сондықтан жарықтандырылған беткі қабатта өмір сүру керек (деп аталады эйфотикалық аймақ ) ның мұхит, теңіз, көл немесе басқа су қоймасы. Фитопланктонның жартысына жуығы келеді фотосинтездеу қызметі Жерде.[6][7][8] Олардың энергияны жинақтауы көміртекті қосылыстар (алғашқы өндіріс ) мұхиттық басым көпшілігінің негізі, сонымен қатар көптеген тұщы су азық-түлік торлары (химосинтез ерекше ерекшелік болып табылады).
Фитопланктонның барлығы дерлік түрлері болып табылады міндетті фотоавтотрофтар, кейбіреулері бар миксотрофты және басқа, пигментті емес түрлері бұл шын мәнінде гетеротрофты (соңғысы көбінесе ретінде қарастырылады зоопланктон ). Олардың ішіндегі ең танымал болып табылады динофлагеллат тұқымдас сияқты Noctiluca және Динофиз, олар алады органикалық көміртегі арқылы ішке қабылдау басқа организмдер немесе детриталь материал.

Фитопланктон тіршілік етеді фотикалық аймақ мұхиттың, қайда фотосинтез мүмкін. Фотосинтез кезінде олар көмірқышқыл газын сіңіріп, оттегін бөліп шығарады. Егер күн радиациясы тым жоғары болса, фитопланктон оның құрбанына айналуы мүмкін фотодеградация. Өсу үшін фитопланктон клеткалары мұхитқа өзендермен, континентальды ауа райымен және полюстердегі мұзды еріген сулармен енетін қоректік заттарға тәуелді. Фитопланктонның бөлінуі еріген органикалық көміртегі (DOC) мұхитқа. Фитопланктон негізі болғандықтан теңіз желілері, олар олжа ретінде қызмет етеді зоопланктон, балық дернәсілдері және басқа да гетеротрофты организмдер. Олар бактериялардың әсерінен немесе әсерінен бұзылуы мүмкін вирустық лизис. Сияқты кейбір фитопланктон жасушалары, дегенмен динофлагеллаттар, тігінен көшуге қабілетті, олар әлі де ағымдарға қарсы белсенді қозғалуға қабілетсіз, сондықтан олар баяу батып, ақыр соңында теңіз қабатын өлі жасушалармен ұрықтандырады және детрит.[9]
Фитопланктон өте маңызды минералдар. Бұл бірінші кезекте макроэлементтер сияқты нитрат, фосфат немесе кремний қышқылы, оның қол жетімділігі деп аталатындар арасындағы тепе-теңдікпен реттеледі биологиялық сорғы және көтерілу қоректік заттарға бай терең сулар. Фитопланктон қоректік құрамы қозғалады және қозғалады Redfield коэффициенті жалпы мұхиттың барлық жерінде болатын макроэлементтер. Алайда мұхиттардың үлкен аудандарында Оңтүстік мұхит, фитопланктон жетіспеушілігімен шектеледі микроэлементтер темір. Бұл кейбір ғалымдардың жақтауына себеп болды темірді ұрықтандыру жиналуына қарсы құрал ретінде адам өндіреді көмірқышқыл газы (СО)2) ішінде атмосфера.[10] Үлкен масштабтағы тәжірибелерде темір қосылды (әдетте, мысалы, тұздар сияқты) темір сульфаты ) фитопланктонның өсуіне ықпал ету және сурет салу үшін мұхиттарға атмосфералық CO2 мұхитқа. Экожүйені манипуляциялау және темірді ұрықтандыру тиімділігі туралы пікірталастар мұндай тәжірибелерді бәсеңдетті.[11]
Фитопланктон тәуелді B дәрумендері өмір сүру үшін. Мұхиттағы аудандарда кейбір В дәрумендерінің, сәйкесінше фитопланктонның жетіспейтіндігі анықталды.[12]
Әсерлері антропогендік жылыну Фитопланктонның ғаламдық популяциясы туралы белсенді зерттеулердің бағыты болып табылады. Су бағанының тік стратификациясының өзгеруі, температураға тәуелді биологиялық реакциялардың жылдамдығы және атмосфералық қоректік заттармен қамтамасыз етілуі болашақ фитопланктон өнімділігіне маңызды әсер етеді деп күтілуде.[13][14]
Мұхиттың антропогендік қышқылдануының фитопланктонның өсуіне және қауымдастық құрылымына әсері де үлкен назар аударды. Кокколитофор сияқты фитопланктон құрамында мұхиттың қышқылдануына сезімтал кальций карбонаты бар жасуша қабырғалары бар. Ұзақ буындардың пайда болуына байланысты, кейбір фитопланктондар жылдамдық шкаласында (айлардан жылдарға дейін) көмірқышқыл газының жоғарылауымен рН өзгеруіне бейімделе алады.[15][16]
Фитопланктон суда өмір сүру үшін маңызды экологиялық функцияны қамтамасыз ете отырып, су қоректену торының негізі ретінде қызмет етеді. Болашақ антропогендік жылыну және мұхиттың қышқылдануы жағдайында фитопланктон өлімінің өзгеруі жылдамдықтың өзгеруіне байланысты зоопланктон жайылым маңызды болуы мүмкін.[17] Көптің бірі тамақ тізбектері мұхитта - сілтемелердің аздығына байланысты керемет - бұл фитопланктонды қолдау крилл (а шаянтәрізділер ұсақ асшаянға ұқсас), ол өз кезегінде қолдайды кит киттер.
Әртүрлілік


Фитопланктон термині судағы барлық фотоавтотрофты микроорганизмдерді қамтиды азық-түлік торлары. Алайда, жердегіден айырмашылығы қауымдастықтар, мұнда автотрофтардың көпшілігі орналасқан өсімдіктер, фитопланктон құрамына кіретін әр түрлі топ протистан эукариоттар және екеуі де эубактериалды және архебактериалды прокариоттар. Теңіз фитопланктонының 5000-ға жуық түрі белгілі.[18] Мұндай әртүрлілік дамыды тапшы ресурстарға қарамастан (шектеу тауашалық саралау ) түсініксіз.[19]
Сандар жағынан фитопланктонның маңызды топтарына мыналар жатады диатомдар, цианобактериялар және динофлагеллаттар, дегенмен көптеген басқа топтар балдырлар ұсынылған. Бір топ кокколитофоридтер, айтарлықтай мөлшерін шығаруға жауапты (ішінара) диметилсульфид (DMS) ішіне атмосфера. DMS бұл тотыққан қоршаған ортадағы сульфатты қалыптастыру аэрозоль бөлшектердің концентрациясы төмен, халықтың санына ықпал ете алады бұлтты конденсация ядролары көбінесе бұлт пен бұлттың көбеюіне әкеледі альбедо сәйкес аталатын CLAW гипотезасы.[20][21] Фитопланктонның әртүрлі түрлері әр түрлі трофикалық деңгейлер әр түрлі экожүйелер шегінде. Жылы олиготрофты сияқты мұхиттық аймақтар Саргассо теңізі немесе Оңтүстік Тынық мұхиты гиры, фитопланктон деп аталатын кішкентай өлшемді жасушалар басым пикопланктон және нанопланктон (пикофлагеллаттар және нанофлагеллаттар деп те аталады), негізінен цианобактериялар (Прохлорококк, Синехококк ) және пикоеукариоттар сияқты Микромоналар. Экожүйелердің басым бөлігі басым көтерілу немесе үлкен жердегі кірістер, үлкенірек динофлагеллаттар фитопланктонның басым бөлігі болып табылады және олардың үлкен бөлігін көрсетеді биомасса.[22]
Өсу стратегиялары
ХХ ғасырдың басында, Альфред С. Редфилд фитопланктонның элементарлы құрамының мұхит тереңіндегі негізгі еріген қоректік заттарға ұқсастығын тапты.[23] Редфилд мұхиттағы көміртектің азот пен фосфорға қатынасы (106: 16: 1) фитопланктонның талаптары бойынша бақыланады деп ұсынды, өйткені фитопланктон кейіннен азот пен фосфорды қайта еске түсіреді. Бұл «Redfield коэффициенті »Сипаттауда стехиометрия Фитопланктон мен теңіз суы теңіз экологиясын, биогеохимияны және фитопланктон эволюциясын түсінудің негізгі принципіне айналды.[24] Алайда, Редфилд коэффициенті әмбебап мән емес және ол экзогендік қоректік заттардың берілуінің өзгеруіне байланысты әр түрлі болуы мүмкін[25] сияқты мұхиттағы микробтық метаболизмдер азотты бекіту, денитрификация және анамокс.
Бір клеткалы балдырларда көрсетілген динамикалық стехиометрия олардың қоректік заттарды ішкі бассейнде сақтау, қоректік заттарға әр түрлі қажеттіліктері бар ферменттер арасында ауысу және осмолиттің құрамын өзгерту қабілетін көрсетеді.[26][27] Әр түрлі жасушалық компоненттердің өзіндік стехиометрия сипаттамалары бар,[24] мысалы, ақуыздар мен хлорофилл сияқты ресурстарды (жеңіл немесе қоректік заттар) жинау машиналарында азоттың мөлшері көп, бірақ фосфоры аз. Сонымен қатар, рибосомалық РНҚ сияқты өсу техникасы құрамында азот пен фосфордың жоғары концентрациясы бар.
Ресурстарды бөлу негізінде фитопланктон өсудің үш түрлі стратегиясына жіктеледі, яғни эквиваленттік, блумер[28] және жалпылама. Survivalist фитопланктонының N: P (> 30) коэффициенті жоғары және аз ресурстардың өсуін қамтамасыз ету үшін ресурстарды жинау техникасының көптігін қамтиды. Блумер фитопланктонының N: P коэффициенті төмен (<10), өсу техникасының үлкен үлесін қамтиды және экспоненциалды өсуге бейімделген. Генералистік фитопланктон N: P-ге және Redfield коэффициентіне ұқсас және ресурстарды алу мен өсірудің салыстырмалы түрде тең техникасын қамтиды.
Молшылыққа әсер ететін факторлар
The NAAMES зерттеуі ғалымдары 2015 пен 2019 жылдар аралығында өткізген бес жылдық ғылыми зерттеу бағдарламасы болды Орегон мемлекеттік университеті және НАСА мұхит экожүйелеріндегі фитопланктон динамикасының аспектілерін және мұндай динамиканың қалай әсер ететінін зерттеу атмосфералық аэрозольдер, бұлттар және климат (NAAMES - Солтүстік Атлантикалық аэрозольдер мен теңіз экожүйелерін зерттеу). Зерттеу барысында Солтүстік Атлант мұхитының субарктикалық аймағына назар аударылды, ол жердегі ең үлкен қайталанатын фитопланктондардың бірі болып табылады. Осы жерде жүргізілген зерттеулердің ұзақ тарихы және қол жетімділіктің салыстырмалы жеңілдігі Солтүстік Атлантиканы басым ғылыми гипотезаларды тексеру үшін тамаша орынға айналдырды.[29] фитопланктонды аэрозоль шығарындыларының Жердің энергетикалық бюджетіндегі рөлін жақсы түсіну мақсатында.[30]
NAAMES жыл сайынғы фитопланктон циклінің нақты фазаларына бағытталған: минимум, шарықтау шегі және биомассаның азаюы мен артуы, бұл гүлденудің пайда болу уақыты мен жыл сайынғы гүлденуді қалпына келтіретін заңдылықтар туралы пікірталастарды шешу үшін.[30] NAAMES жобасы сонымен бірге өндірілген аэрозольдердің мөлшерін, мөлшерін және құрамын зерттеді алғашқы өндіріс фитопланктонның гүлдену циклдарының бұлт түзілімдері мен климатқа қалай әсер ететінін түсіну үшін.[31]





Фитопланктонның рөлі

Оң жақтағы диаграммада фитопланктон әсер ететін бөлімдерге атмосфералық газ құрамы, бейорганикалық қоректік заттар және микроэлементтер ағындары, сондай-ақ биологиялық процестер арқылы органикалық заттардың ауысуы мен циклі жатады. Фотосинтезделген тіркелген көміртек тез қайта өңделеді және жер үсті мұхитында қайта пайдаланылады, ал бұл биомассаның белгілі бір бөлігі батып бара жатқан бөлшектер ретінде терең мұхитқа экспортталады, ол трансформация процестеріне ұшырайды, мысалы, қайта қалпына келтіру.[35]
Аквамәдениет
Фитопланктон - екеуінде де негізгі тағамдық зат аквамәдениет және марикультура. Екеуі де фитопланктонды өсірілетін жануарларға тамақ ретінде пайдаланады. Марикультурада фитопланктон табиғи түрде кездеседі және теңіз суының қалыпты айналымымен қоршауларға енгізіледі. Аквамәдениетте фитопланктонды тікелей алып, енгізу керек. Планктонды су айдынынан жинауға немесе өсіруге болады, дегенмен бұрынғы әдіс сирек қолданылады. Фитопланктон өндіріс үшін тағамдық шикізат ретінде қолданылады айналдырғыштар,[36] олар өз кезегінде басқа организмдерді тамақтандыру үшін қолданылады. Фитопланктон аквамәдениеттің көптеген сорттарын тамақтандыру үшін де қолданылады моллюскалар, оның ішінде інжу устрицалар және алып моллюскалар. 2018 жылғы зерттеу табиғи фитопланктонның қоректік құндылығын көмірсулар, протеиндер және әлемдік мұхиттағы липидтер тұрғысынан спутниктерден алынған мұхит түсі мәліметтерін бағалады,[37] Фитопланктонның калориялық мәні әр түрлі мұхиттық аймақтарда және жылдың әр түрлі уақытында әр түрлі болатындығын анықтады.[37][38]
Жасанды жағдайда фитопланктон өндірісінің өзі аквамәдениеттің бір түрі болып табылады. Фитопланктон әртүрлі мақсаттарда өсіріледі, соның ішінде басқа аквамәдени организмдерге арналған тамақ өнімдері,[36] тұтқындауға арналған тағамдық қоспалар омыртқасыздар жылы аквариум. Мәдениеттің өлшемдері кішігірім масштабта болады зертхана коммерциялық аквамәдениет үшін 1L-ден бірнеше ондаған мың литрге дейінгі дақылдар.[36] Мәдениеттің көлеміне қарамастан, планктондардың тиімді өсуі үшін белгілі бір жағдайлар жасалуы керек. Мәдениетті планктондардың көпшілігі теңіз және теңіз суы а меншікті салмақ Қоректік орта ретінде 1,010-дан 1,026-ға дейін қолдануға болады. Бұл су болуы керек зарарсыздандырылған, әдетте жоғары температурада автоклав немесе әсер ету арқылы ультрафиолет сәулеленуі, алдын алу биологиялық ластану мәдениет. Әр түрлі тыңайтқыштар планктонның өсуін жеңілдету үшін қоректік ортаға қосылады. Планктонды тоқтата тұру үшін, сондай-ақ ерігенді қамтамасыз ету үшін дақыл қандай-да бір жолмен газдалуы немесе қозуы керек Көмір қышқыл газы үшін фотосинтез. Тұрақты аэрациядан басқа, көптеген дақылдар қолмен араластырылады немесе тұрақты түрде араластырылады. Фитопланктонның өсуі үшін жарық беру керек. The түс температурасы жарықтандыру шамамен 6500 К болуы керек, бірақ 4000 К-ден 20000 К жоғары мәндер сәтті қолданылды. Жарық сәулесінің ұзақтығы күніне шамамен 16 сағат болуы керек; бұл ең тиімді жасанды күн ұзақтығы.[36]
Антропогендік өзгерістер
Теңіз фитопланктоны ғаламдық фотосинтетикалық СО-ның жартысын орындайды2 бекіту (жылына ~ 50 Pg C таза ғаламдық бастапқы өндіріс) және оттегінің жартысы өсімдіктердің әлемдік биомассасының ~ 1% -ына тең болғанымен.[39] Құрлықтағы өсімдіктермен салыстырғанда теңіз фитопланктоны үлкен беткейге таралған, аз маусымдық өзгеріске ұшырайды және ағаштармен салыстырғанда жылдам айналым жылдамдығына ие (онжылдықтармен салыстырғанда күндер).[39] Сондықтан фитопланктон климаттың өзгеруіне ғаламдық ауқымда тез жауап береді. Бұл сипаттамалар фитопланктонның көміртекті бекітуге қосқан үлесін бағалап, бұл өндіріс тербелістерге байланысты қалай өзгеруі мүмкін екенін болжау кезінде маңызды. Климаттың өзгеруінің алғашқы өнімділікке әсерін болжау фитопланктонның гүлдену циклдарымен қиындатылады, оларға төменнен бақылау (мысалы, маңызды қоректік заттардың болуы және вертикалды араластыру) және жоғарыдан төмен бақылау (мысалы, жайылым және вирус) әсер етеді.[40][39][41][42][43][44] Күн радиациясының, температураның және тұщы сулардың жер үсті суларына түсуінің артуы мұхиттың стратификациясын күшейтеді және соның салдарынан қоректік заттардың терең судан жер үсті суларына өтуін азайтады, бұл алғашқы өнімділікті төмендетеді.[39][44][45] Керісінше, CO-ның жоғарылауы2 деңгейлері фитопланктонның алғашқы өндірісін арттыра алады, бірақ қоректік заттар шектеусіз болған кезде ғана.[46][47][48][17]
Кейбір зерттеулер өткен ғасырда жалпы әлемдік фитопланктонның тығыздығы төмендегенін көрсетеді,[49] бірақ бұл тұжырымдар ұзақ мерзімді фитопланктон мәліметтерінің қол жетімділігі шектеулі болғандықтан, деректерді қалыптастырудағы әдіснамалық айырмашылықтар және фитопланктон өндірісіндегі үлкен жылдық және декадалық өзгергіштіктер болды.[50][51][52][53] Сонымен қатар, басқа зерттеулер мұхиттық фитопланктон өндірісінің жаһандық өсуін ұсынады[54] және белгілі бір аймақтардағы немесе белгілі бір фитопланктон топтарындағы өзгерістер.[55][56] Әлемдік теңіз мұзының индексі төмендеуде,[57] жарықтың жоғары енуіне және бастапқы өндіріске ықтимал көбірек әкелуге;[58] дегенмен, ауыспалы араластыру заңдылықтары мен қоректік заттардың өзгеруінің әсерлері және полярлық аймақтардағы өнімділік тенденциясы туралы қарама-қайшы болжамдар бар.[44][17]
Сондай-ақ қараңыз
- Балдырлар - фотосинтетикалық эукариоттық организмдердің әр түрлі тобы
- Алгультура - балдырлар өсіруді қамтитын аквамәдениет
- Балдырлар негізі
- Бактериопланктон - су бағанында қозғалатын планктонның бактериалды компоненті
- Биологиялық сорғы - Мұхиттың атмосферадан мұхиттың ішкі бөлігі мен теңіз қабатына дейінгі биологиялық қозғалатын секвестрі
- CLAW гипотезасы - теңіз биотасы мен климатты байланыстыратын болжамды теріс кері байланыс
- Цианобактериялар - Прокариоттардың фотосинтездеу филумы
- Тұщы су фитопланктоны
- Темірді ұрықтандыру
- Микрофит (микробалдырлар)
- NAMAMES
- Мұхиттың қышқылдануы - көміртегі диоксидінің түсуіне байланысты Жер мұхитының рН-нің тұрақты төмендеуі
- Планктон парадоксы - Аз ресурстарға бәсекелестікке қарамастан, планктондардың әртүрлілігін экологиялық бақылау
- Фотосинтетикалық пикопланктон
- Планктон - су бағанында және ағысқа қарсы жүзе алмайтын ағзалар
- Whiting іс-шарасы - су қоймаларындағы ұсақ түйіршікті кальций карбонат бөлшектерінің суспензиясы
- Зоопланктон - планетон экожүйесінің гетеротрофты протистаны немесе метазоа мүшелері
- Жіңішке қабаттар (океанография)
- Зиянды балдырлар гүлдейді - Ағзалардың популяциялық жарылуы (әдетте балдырлар ) табиғи теңіздегі оттегінің деңгейін айтарлықтай төмендетіп, теңіз тіршілігін жоя алады
Әдебиеттер тізімі
- ^ Турман, H. V. (2007). Кіріспе океанография. Академиялық Интернет баспагерлері. ISBN 978-1-4288-3314-2.[бет қажет ]
- ^ Ghosal; Роджерс; Рей, С .; М .; А. «Турбуленттіліктің фитопланктонға әсері». Аэроғарыштық технологиялар кәсіпорны. NTRS. Алынған 16 маусым 2011.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ а б Жаһандық мұхиттағы фитопланктондық қауымдастықтар NASA Hyperwall, 30 қыркүйек 2015 ж.
 Бұл мақалада осы қайнар көздегі мәтін енгізілген қоғамдық домен.
Бұл мақалада осы қайнар көздегі мәтін енгізілген қоғамдық домен. - ^ Линдси, Р., Скотт, М. және Симмон, Р. (2010) «Фитопланктон дегеніміз не». NASA Жер обсерваториясы.
- ^ Дарвин жобасы Массачусетс технологиялық институты.
- ^ Майкл Дж.Беренфельд; т.б. (30 наурыз 2001). «ENSO ауысуы кезіндегі биосфералық алғашқы өндіріс» (PDF). Ғылым. 291 (5513): 2594–7. Бибкод:2001Sci ... 291.2594B. дои:10.1126 / ғылым.1055071. PMID 11283369. S2CID 38043167.
- ^ «NASA жер серігі ғаламдық мұхит өсімдіктерінің денсаулығын сақтаудың қызыл сәулесін анықтады» НАСА, 28 мамыр 2009 ж.
- ^ «Спутниктік мұхит өсімдіктері көбейеді, жасылдануды жағалайды». НАСА. 2 наурыз 2005 ж. Алынған 9 маусым 2014.
- ^ а б Käse L, Geuer JK. (2018) «Теңіз климатының өзгеруіне фитопланктон реакциясы - кіріспе». Jungblut S., Liebich V., Bode M. (Eds) YOUMARES 8 –Шекаралар арқылы мұхиттар: бір-бірінен үйрену, 55-72 беттер, Шпрингер. дои:10.1007/978-3-319-93284-2_5.
 Материал осы дереккөзден көшірілген, ол а Creative Commons Attribution 4.0 Халықаралық лицензиясы.
Материал осы дереккөзден көшірілген, ол а Creative Commons Attribution 4.0 Халықаралық лицензиясы. - ^ Richtel, M. (1 мамыр 2007). «Жаһандық жылынумен күресу үшін планктон тарту». New York Times.
- ^ Монастерский, Ричард (1995). «Темір парникке қарсы: Мұхиттанушылар ғаламдық жылыну терапиясын абайлап зерттейді». Ғылым жаңалықтары. 148 (14): 220–1. дои:10.2307/4018225. JSTOR 4018225.
- ^ Саньудо-Вильгельми, Серхио (23 маусым 2012). «Мұхитта» шөлдер «витаминінің болуы расталды». ScienceDaily.
- ^ Хенсон, С.А .; Сармиенто, Дж. Л .; Данн, Дж. П .; Бопп, Л .; Лима, Мен .; Дони, С. С .; Джон Дж .; Beauleu, C. (2010). «Мұхит хлорофиллінің және өнімділіктің спутниктік жазбаларында климаттың антропогендік өзгеруін анықтау». Биогеология. 7 (2): 621–40. Бибкод:2010BGeo .... 7..621H. дои:10.5194 / bg-7-621-2010.
- ^ Штайнахер, М .; Джус, Ф .; Фроличер, Т.Л .; Бопп, Л .; Кадул, П .; Кокко, V .; Дони, С. С .; Гехлен М .; Линдсей, К .; Мур, Дж. К .; Шнайдер, Б .; Segschneider, J. (2010). «Теңіз өнімділігінің болжамды ХХІ ғасырдағы төмендеуі: көп модельді талдау». Биогеология. 7 (3): 979–1005. Бибкод:2010BGeo .... 7..979S. дои:10.5194 / bg-7-979-2010.
- ^ Коллинз, Синеад; Рост, Бьорн; Райнирсон, Татьяна А. (25 қараша 2013). «Мұхитты қышқылдандыру кезіндегі теңіз фитопланктонының эволюциялық әлеуеті». Эволюциялық қосымшалар. 7 (1): 140–155. дои:10.1111 / eva.12120. ISSN 1752-4571. PMC 3894903. PMID 24454553.
- ^ Лохбек, Кай Т .; Рибеселл, Ульф; Reusch, Thorsten B. H. (8 сәуір 2012). «Фитопланктонның негізгі түрлерінің мұхиттың қышқылдануына бейімделу эволюциясы». Табиғи геология. 5 (5): 346–351. Бибкод:2012NatGe ... 5..346L. дои:10.1038 / ngeo1441. ISSN 1752-0894.
- ^ а б c Кавиччиоли, Рикардо; Риппл, Уильям Дж.; Тиммис, Кеннет Н .; Азам, Фарук; Бакен, Ларс Р .; Байлис, Матай; Бехренфельд, Майкл Дж .; Боетиус, Антье; Бойд, Филипп В. Classen, Aimée T .; Кротер, Томас В.; Дановаро, Роберто; Бригадир, Кристин М .; Хуисман, Джеф; Хатчинс, Дэвид А .; Янссон, Джанет К .; Карл, Дэвид М .; Коскелла, Бритт; Марк Уэлч, Дэвид Б .; Мартини, Дженнифер Б. Х .; Моран, Мэри Анн; Жетім, Виктория Дж.; Рэй, Дэвид С .; Ремаис, Джастин V .; Рич, Вирджиния I .; Сингх, Браджеш К .; Стейн, Лиза Ю .; Стюарт, Фрэнк Дж .; Салливан, Мэттью Б .; т.б. (2019). «Ғалымдардың адамзатқа ескертуі: микроорганизмдер және климаттың өзгеруі». Микробиологияның табиғаты туралы шолулар. 17 (9): 569–586. дои:10.1038 / s41579-019-0222-5. PMC 7136171. PMID 31213707. Материал осы дереккөзден көшірілген, ол а Creative Commons Attribution 4.0 Халықаралық лицензиясы.
- ^ Халлеграеф, Г.М. (2003). «Зиянды балдырлар гүлдейді: ғаламдық шолу» (PDF). Галлеграефте Густааф М .; Андерсон, Дональд Марк; Цембелла, Аллан Д .; Эневольдсен, Генрик О. (ред.) Зиянды теңіз микробалдырлары жөніндегі нұсқаулық. Юнеско. 25-49 бет. ISBN 978-92-3-103871-6.
- ^ Хатчинсон, Г.Э. (1961). «Планктон парадоксы». Американдық натуралист. 95 (882): 137–45. дои:10.1086/282171. S2CID 86353285.
- ^ Чарлсон, Роберт Дж.; Ловлок, Джеймс Э .; Андреа, Мейнрат О .; Уоррен, Стивен Г. (1987). «Мұхиттық фитопланктон, атмосфералық күкірт, бұлтты альбедо және климат». Табиғат. 326 (6114): 655–61. Бибкод:1987 ж. 326..655С. дои:10.1038 / 326655a0. S2CID 4321239.
- ^ Куинн, П. К .; Бейтс, Т.С (2011). «Мұхиттық фитопланктон күкірт шығарындылары арқылы климатты реттеуге қарсы іс». Табиғат. 480 (7375): 51–6. Бибкод:2011 ж. 480 ... 51Q. дои:10.1038 / табиғат 1055. PMID 22129724. S2CID 4417436.
- ^ Калбет, А. (2008). «Теңіз жүйелеріндегі микрозоопланктонның трофикалық рөлдері». ICES журналы теңіз ғылымы. 65 (3): 325–31. дои:10.1093 / icesjms / fsn013.
- ^ Редфилд, Альфред С. (1934). «Теңіз суындағы органикалық туындылардың пропорциясы және олардың планктон құрамымен байланысы туралы». Джонстоун, Джеймс; Даниэль, Ричард Джеллико (ред.) Джеймс Джонстонның мемориалдық томы. Ливерпуль: University Liver of Liverpool. 176–92 бет. OCLC 13993674.
- ^ а б Арриго, Кевин Р. (2005). «Теңіз микроорганизмдері және ғаламдық қоректік циклдар». Табиғат. 437 (7057): 349–55. Бибкод:2005 ж.437..349А. дои:10.1038 / табиғат04159. PMID 16163345. S2CID 62781480.
- ^ Фэннинг, Кент А. (1989). «Мұхиттағы қоректік заттардың шектелуіне атмосфераның ластануының әсері». Табиғат. 339 (6224): 460–63. Бибкод:1989 ж.33..460F. дои:10.1038 / 339460a0. S2CID 4247689.
- ^ Штернер, Роберт Уорнер; Элсер, Джеймс Дж. (2002). Экологиялық стоихиометрия: Молекулалардан биосфераға дейінгі элементтер биологиясы. Принстон университетінің баспасы. ISBN 978-0-691-07491-7.[бет қажет ]
- ^ Клаусмейер, Кристофер А .; Литчман, Елена; Левин, Саймон А. (2004). «Фитопланктонның өсуі және қоректік заттардың бірнеше шектелуіндегі стехиометрия». Лимнология және океанография. 49 (4 2 бөлім): 1463–70. Бибкод:2004LimOc..49.1463K. дои:10.4319 / lo.2004.49.4_part_2.1463. S2CID 16438669.
- ^ Клаусмейер, Кристофер А .; Литчман, Елена; Дауфресне, Тангуй; Левин, Саймон А. (2004). «Фитопланктонның азот-фосфор стехиометриясы оңтайлы». Табиғат. 429 (6988): 171–4. Бибкод:2004 ж. Табиғат.429..171K. дои:10.1038 / табиғат02454. PMID 15141209. S2CID 4308845.
- ^ а б Бехренфельд, МДж және Босс, Е.С. (2018 ж.) «Фитопланктонның жылдық циклдері аясында гүлдену гипотезалары бойынша студенттерге арналған оқу құралы». Ғаламдық өзгерістер биологиясы, 24(1): 55–77. дои:10.1111 / gcb.13858.
- ^ а б Бехренфельд, Майкл Дж .; Мур, Ричард Х .; Хостетлер, Крис А .; Графф, Джейсон; Гаубе, Питер; Рассел, Линн М .; Чен, Гао; Дони, Скотт С .; Джованнони, Стивен; Лю, Хуню; Проктор, Кристофер (22 наурыз 2019). «Солтүстік Атлантикалық аэрозоль мен теңіз экожүйесін зерттеу (NAAMES): ғылыми мотив және миссияға шолу». Теңіз ғылымындағы шекаралар. 6: 122. дои:10.3389 / fmars.2019.00122. ISSN 2296-7745.
- ^ Энгель, Аня; Банж, Герман В .; Кунлифф, Майкл; Берроуз, Сюзанна М .; Фридрихс, Герно; Галгани, Луиза; Герман, Хартмут; Херткорн, Норберт; Джонсон, Мартин; Лисс, Питер С.; Куинн, Патриция К. (30 мамыр 2017). «Мұхиттың маңызды терісі: теңіз бетіндегі микроқабатты интеграцияланған түсінуге бағытталған». Теңіз ғылымындағы шекаралар. 4. дои:10.3389 / fmars.2017.00165. ISSN 2296-7745.
- ^ Бехренфельд, Майкл Дж .; Босс, Эммануил С. (3 қаңтар 2014). «Мұхит планктонының гүлденуінің экологиялық негіздерін қайта құру». Жыл сайынғы теңіз ғылымына шолу. 6 (1): 167–194. Бибкод:2014ARMS .... 6..167B. дои:10.1146 / annurev-marine-052913-021325. ISSN 1941-1405. PMID 24079309. S2CID 12903662.
- ^ NAAMES: Ғылым - мақсаттар Лэнгли зерттеу орталығы, NASA, Жаңартылған: 6 маусым 2020. Шығарылды: 15 маусым 2020.
- ^ а б Ригетти, Д., Фогт, М., Грубер, Н., Псомас, А. және Циммерманн, Н.Е. (2019) «Температура мен қоршаған ортаның өзгермелілігі әсер ететін фитопланктонның әртүрлілігінің ғаламдық үлгісі». Ғылым жетістіктері, 5(5): eaau6253. дои:10.1126 / sciadv.aau6253. Материал осы дереккөзден көшірілген, ол а Creative Commons Attribution 4.0 Халықаралық лицензиясы.
- ^ а б Генрихс, Мара Е .; Мори, Коринна; Длюгощ, Леон (2020). «Су ағзалары мен олардың әртүрлі көзқарастар бойынша анықталған химиялық ортасының арасындағы күрделі өзара әрекеттесу». YOUMARES 9 - Мұхиттар: біздің зерттеулеріміз, біздің болашағымыз. 279–297 беттер. дои:10.1007/978-3-030-20389-4_15. ISBN 978-3-030-20388-7. Материал осы дереккөзден көшірілген, ол а Creative Commons Attribution 4.0 Халықаралық лицензиясы.
- ^ а б c г. Макви, Джеймс П., Най-Сянь Чао және Ченг-Шенг Ли. CRC Mariculture анықтамалығы том. 1: шаян тәрізділерді өсіру. Нью-Йорк: C R C P LLC, 1993 ж.[бет қажет ]
- ^ а б Рой, Шовонлал (12 ақпан 2018). «Фитопланктон көмірсуларының, ақуыздың және липидтің әлемдік мұхиттағы спутниктік мұхит түсінен таралуы». ISME журналы. 12 (6): 1457–1472. дои:10.1038 / s41396-018-0054-8. ISSN 1751-7370. PMC 5955997. PMID 29434313.
- ^ «Тамақтануды зерттеу әлемдегі балық аулаудың маңызды аймақтарындағы тұрақсыздықты анықтады».
- ^ а б c г. Бехренфельд, Майкл Дж. (2014). «Планктонның климаттық биі». Табиғи климаттың өзгеруі. 4 (10): 880–887. Бибкод:2014 ж. NATCC ... 4..880B. дои:10.1038 / nclimate2349.
- ^ Хатчинс, Д. А .; Бойд, П.В. (2016). «Теңіз фитопланктоны және өзгеріп жатқан мұхит темір циклі». Табиғи климаттың өзгеруі. 6 (12): 1072–1079. Бибкод:2016NatCC ... 6.1072H. дои:10.1038 / nclimate3147.
- ^ Де Баар, Хейн Дж. В .; Де Йонг, Джерун Т.М .; Баккер, Доротье С. Е .; Лёшер, Беттина М .; Вет, Корнелис; Батманн, Ули; Сметачек, Виктор (1995). «Оңтүстік мұхитта темірдің планктондардың гүлденуі мен көмірқышқыл газының азаюы үшін маңызы». Табиғат. 373 (6513): 412–415. Бибкод:1995 ж.33..412D. дои:10.1038 / 373412a0. S2CID 4257465.
- ^ Бойд, П.В .; Джикеллс, Т .; Заң, C. S .; Блейн, С .; Бойль, Э. А .; Бюселер, К.О .; Коул, К. Х .; Каллен, Дж. Дж .; Де Баар, H. J. W .; Іздейді, М .; Харви, М .; Ланселот, С .; Левасир М .; Оуэнс, N. P. J .; Поллард, Р .; Ривкин, Р.Б .; Сармиенто, Дж .; Шоеманн, V .; Сметачек, V .; Такеда, С .; Цуда, А .; Тернер, С .; Уотсон, Дж. (2007). «1993-2005 жж. Темірді байыту бойынша тәжірибе: синтез және болашақ бағыттары» (PDF). Ғылым. 315 (5812): 612–617. Бибкод:2007Sci ... 315..612B. дои:10.1126 / ғылым.1131669. PMID 17272712. S2CID 2476669.
- ^ Бехренфельд, Майкл Дж .; оМэлли, Роберт Т .; Босс, Эммануэль С .; Вестберри, Тоби К .; Графф, Джейсон Р .; Хэлси, Кимберли Х .; Миллиган, Аллен Дж .; Зигель, Дэвид А .; Браун, Мэтью Б. (2016). «Мұхит жылынуының жаһандық фитопланктонға әсерін қайта бағалау». Табиғи климаттың өзгеруі. 6 (3): 323–330. Бибкод:2016NatCC ... 6..323B. дои:10.1038 / nclimate2838.
- ^ а б c Бехренфельд, Майкл Дж .; Ху, Юнсян; оМэлли, Роберт Т .; Босс, Эммануэль С .; Хостетлер, Крис А .; Зигель, Дэвид А .; Сармиенто, Хорхе Л. Шулиен, Дженнифер; Шаш, Джонатан В.; Лу, Сяомей; Родье, Шарон; Скарино, Эми Джо (2017). «Полярлы фитопланктон биомассасының жыл сайынғы серпіліс-циклдары ғарыштық лидармен анықталды». Табиғи геология. 10 (2): 118–122. Бибкод:2017NatGe..10..118B. дои:10.1038 / ngeo2861.
- ^ Бехренфельд, Майкл Дж .; оМэлли, Роберт Т .; Зигель, Дэвид А .; Макклейн, Чарльз Р .; Сармиенто, Хорхе Л. Фельдман, Джин С .; Миллиган, Аллен Дж .; Фальковский, Пол Г .; Летелье, Рикардо М .; Босс, Эммануил С. (2006). «Мұхиттың қазіргі заманғы өнімділігінің климаттық үрдістері». Табиғат. 444 (7120): 752–755. Бибкод:2006 ж. 4444..752B. дои:10.1038 / табиғат05317. PMID 17151666. S2CID 4414391.
- ^ Левитан, О .; Розенберг, Г .; Сетлик, I .; Сетликова, Е .; Григель, Дж .; Клепетар, Дж .; Прасил, О .; Берман-Франк, И. (2007). «СО2 жоғарылауы азоттың бекітілуін және теңіз цианобактериясының Trichodesmium өсуін күшейтеді». Ғаламдық өзгерістер биологиясы. 13 (2): 531–538. Бибкод:2007GCBio..13..531L. дои:10.1111 / j.1365-2486.2006.01314.x.
- ^ Верспаген, Жоланда М. Х .; Ван Де Ваал, Дедмер Б .; Финке, Ян Ф .; Виссер, Петра М .; Хуисман, Джеф (2014). «СО2 жоғарылауының алғашқы өндіріске және экологиялық стехиометрияға әртүрлі қоректік деңгейлердегі қарама-қарсы әсерлері» (PDF). Экология хаттары. 17 (8): 951–960. дои:10.1111 / ele.12298. PMID 24813339.
- ^ Холдинг, Дж. М .; Дуарте, К.М .; Санц-Мартин, М .; Меса, Э .; Арриета, Дж. М .; Чиери, М .; Хендрикс, И. Е .; Гарсия-Коррал, Л.С .; Регауди-Де-Джу, А .; Делгадо, А .; Рейгстад, М .; Вассманн, П .; Agustí, S. (2015). «CO температураға тәуелділігі2- Еуропалық Солтүстік Мұзды мұхиттағы алғашқы өндіріс ». Табиғи климаттың өзгеруі. 5 (12): 1079–1082. Бибкод:2015NatCC ... 5.1079H. дои:10.1038 / nclimate2768. hdl:10754/596052.
- ^ Бойс, Даниэль Дж.; Льюис, Марлон Р .; Құрт, Борис (2010). «Өткен ғасырда ғаламдық фитопланктонның құлдырауы». Табиғат. 466 (7306): 591–596. Бибкод:2010 ж. 466..591B. дои:10.1038 / табиғат09268. PMID 20671703. S2CID 2413382.
- ^ МакКас, Дэвид Л. (2011). «Хлорофилл деректерінің араласуы уақытша тенденцияны білдіреді ме?». Табиғат. 472 (7342): E4-E5. Бибкод:2011 ж. 472E ... 4M. дои:10.1038 / nature09951. PMID 21490623. S2CID 4308744.
- ^ Рыкачевский, Райан Р. Данн, Джон П. (2011). «Мұхиттағы хлорофилл трендтеріне өлшенген көзқарас». Табиғат. 472 (7342): E5-E6. Бибкод:2011 ж. 472E ... 5R. дои:10.1038 / табиғат09952. PMID 21490624. S2CID 205224535.
- ^ Мак-Каттарс-Голлоп, Эбигейл; Рейд, Филипп С .; Эдвардс, Мартин; Беркилл, Питер Х.; Кастеллани, Клаудия; Баттен, Соня; Гизкес, Уинфрид; Би, Даг; Бидигаре, Роберт Р .; Басшысы, Эрика; Джонсон, Род; Кахру, Мати; Кослоу, Дж. Энтони; Пена, Анжелика (2011). «Теңіз фитопланктонының төмендеуі бар ма?». Табиғат. 472 (7342): E6-E7. Бибкод:2011 ж. 472E ... 6M. дои:10.1038 / табиғат09950. PMID 21490625. S2CID 205224519.
- ^ Бойс, Даниэл Дж.; Льюис, Марлон Р .; Ворм, Борис (2011). «Бойс және басқалар. Жауап беру». Табиғат. 472 (7342): E8-E9. Бибкод:2011 ж. 472E ... 8B. дои:10.1038 / табиғат09953. S2CID 4317554.
- ^ Антуан, Дэвид (2005). «Ұзақ мерзімді үрдістерді іздеу үшін 1980-ші және 2000-ші жылдардағы мұхиттардың түрлі-түсті бақылауларын көбейту». Геофизикалық зерттеулер журналы. 110 (C6): C06009. Бибкод:2005JGRC..110.6009A. дои:10.1029 / 2004JC002620.
- ^ Уернанд, Марсель Р .; Ван Дер Верд, Хендрик Дж.; Gieskes, Winfried W. C. (2013). «Мұхит түсі мен хлорофилл концентрациясының тенденциялары, 1889-2000 жылдар аралығында, бүкіл әлемде». PLOS ONE. 8 (6): e63766. Бибкод:2013PLoSO ... 863766W. дои:10.1371 / journal.pone.0063766. PMC 3680421. PMID 23776435.
- ^ Руссо, Сесиль С.; Грегг, Уотсон В. (2015). «Жаһандық фитопланктон құрамының соңғы онжылдық тенденциялары». Әлемдік биогеохимиялық циклдар. 29 (10): 1674–1688. Бибкод:2015GBioC..29.1674R. дои:10.1002 / 2015GB005139.
- ^ Теңіз мұзының индексі Ұлттық қар мен мұз туралы мәліметтер орталығы. Қолданылды 30 қазан 2020.
- ^ Кирхман, Дэвид Л .; Моран, Хосе Анхелу Г .; Даклоу, Хью (2009). «Полярлық мұхиттағы микробтық өсу - температураның рөлі және климаттың өзгеруіне әсер етуі». Микробиологияның табиғаты туралы шолулар. 7 (6): 451–459. дои:10.1038 / nrmicro2115. PMID 19421189. S2CID 31230080.
Әрі қарай оқу
- Грисон, Филлип Э. (1982). Америка Құрама Штаттарының Фитопланктонында байқалған балдырлардың жиі кездесетін және басым тұқымдарын анықтауға арналған түсіндірме кілт. Вашингтон, Колумбия округі: Америка Құрама Штаттарының мемлекеттік баспа кеңсесі ISBN 978-0-607-68844-3.
- Кирби, Ричард Р. (2010). Мұхиттың дрейферлері: толқындар астындағы құпия әлем. Кактус студиясы. ISBN 978-1-904239-10-9.
- Мартин, Рональд; Квигг, Антониетта (2013). «Бір кездері теңіздерді басқарған кішкентай өсімдіктер». Ғылыми американдық. 308 (6): 40–5. Бибкод:2013SciAm.308f..40M. дои:10.1038 / Scientificamerican0613-40. PMID 23729069.
Сыртқы сілтемелер
- Secchi Disk және Secchi қосымшасы, фитопланктонды зерттеу бойынша ғылыми жоба
- Ocean Drifters, Дэвид Аттенборо айтқан планктонның әртүрлі рөлдері туралы қысқаша фильм
- Планктон шежіресі, қысқаметражды деректі фильмдер мен фотосуреттер
- DMS және климат, NOAA
- Планктон * Желі, планктоникалық түрлердің суреттері

Су экожүйелері | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||
| |||||||||||||||||


| Билікті бақылау |
|---|