Тамырлы камбий - Vascular cambium
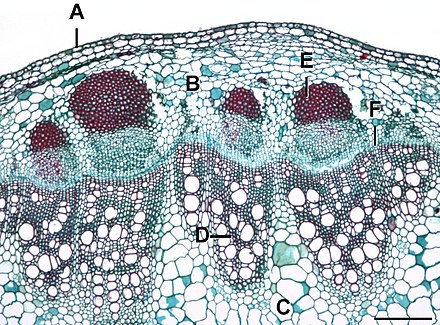
The тамырлы камбий негізгі өсім болып табылады мата көптеген өсімдіктердің сабақтары мен тамырларында, атап айтқанда дикоттар сары май мен емен ағаштары сияқты, гимноспермалар қарағай сияқты, сондай-ақ белгілі бір тамырлы өсімдіктер. Ол екінші ретті шығарады ксилема ішке қарай пит, және екінші флоэма сыртқа қарай қабығы.
Шөптесін өсімдіктерде бұл көбінесе сабақтың ішінде үзілген сақина түзетін алқадағы моншақ тәрізді орналасқан тамырлы буындарда болады. Ағаш өсімдіктерінде ол мамандандырылмаған цилиндр құрайды меристема жасушалар, олардан жаңа ұлпалар өсетін үздіксіз сақина ретінде. Ол ксилема мен флоэмадан айырмашылығы өсімдік арқылы суды, минералды заттарды немесе тағамды тасымалдамайды. Қантамырдың басқа атаулары камбий болып табылады негізгі камбий, ағаш камбийі, немесе екі жақты камбий.
Пайда болу
Тамырлы камбия табылған дикоттар және гимноспермалар бірақ жоқ монокоттар, әдетте оларда екінші өсу болмайды. Бірнеше жапырақ түрлерінде тамырлы камбий болады. Дикотта және гимноспермада ағаштар, тамырлы камбий - бұл қабық пен ағашты бөлетін айқын сызық; оларда да бар тығын камбийі. Сәтті үшін егу, тамыр мен камбияның тамырлы камбиясы бір-біріне сәйкес өсуі үшін бір-біріне сәйкестендірілуі керек.
Құрылымы және қызметі
Бастапқы ксилема мен бастапқы флоэма арасында кездесетін камбий деп аталады intrafascicular камбий (тамыр шоғырларының ішінде). Екінші өсу кезінде медулярлық сәулелердің жасушалары бір сызық бойымен (бөлімде көрсетілгендей; үш өлшемде ол парақ) көршілес тамыр шоғырларының арасында болады. меристемалық және жаңа қалыптастыру интерфасикулярлық камбий (тамырлы шоқтар арасында). Интрасасикулярлық және интерфасикулярлық камбия осылайша біріктіріліп, алғашқы ксилема мен бастапқы флоэманы бөлетін сақина (үш өлшемді түтік) құрайды. камбий сақинасы. Тамырлы камбий сақинаның ішкі жағында екінші реттік ксилема, ал сыртында екінші ретті флоэма түзіп, біріншілік ксилема мен флоэманы алшақтатады.
Тамырлы камбий әдетте екі типті жасушадан тұрады:
- Fusiform инициалдары (биік, осьтік бағытталған)
- Сәуле инициалдары (пішіні кішірек және дөңгелек пішінді)
Камбиальды меристеманы қолдау
Тамырлы камбий өзара әрекеттесетін кері байланыс циклдарының желісімен қамтамасыз етіледі. Қазіргі уақытта гормондар да, қысқа пептидтер де осы жүйелердегі ақпарат тасымалдаушылар ретінде анықталды. Осындай реттеу басқа өсімдіктерде кездеседі меристемалар, камбиальды меристема меристема үшін ксилема жағынан да, флоэма жағынан да сигналдар қабылдайды. Меристемадан тыс сигналдар жасушалардың көбеюіне және дифференциациясына ықпал ететін ішкі факторларды реттейді.[1]
Гормоналды реттеу
The фитогормондар тамырлы камбиальды белсенділікке қатысады ауксиндер, этилен, гиббереллиндер, цитокининдер, абциз қышқылы және, бәлкім, көп нәрсе табылуы мүмкін. Осы өсімдік гормондарының әрқайсысы камбиальды белсенділікті реттеу үшін өте маңызды. Осы гормондардың әр түрлі концентрациясының үйлесуі өсімдіктер алмасуында өте маңызды.
Ауксин гормондарының ынталандыратыны дәлелденген митоз, жасуша өндірісі және интерфасикулярлық және фасцикулярлық камбийді реттейді. Ауксинді ағаш діңінің бетіне жағу бас кесілген өсінділерге екінші өсуді жалғастыруға мүмкіндік берді. Ауксин гормондарының болмауы өсімдікке зиянды әсер етеді. Ауксинсіз мутанттардың интерфасикулярлық камбийлердің аралықтары кеңейіп, өсінділері төмендейтіні көрсетілген. тамырлы шоқтар. Сондықтан мутантты өсімдік бүкіл өсімдік бойымен тасымалданатын судың, қоректік заттардың және фотосинтаттардың азаюын сезінеді, нәтижесінде өлімге әкеледі. Ауксин сонымен қатар тамырлы камбийдегі жасушаның екі түрін, сәулелік және фузиформалы инициалдарды реттейді. Осы инициалдарды реттеу қоректік транслокация үшін ксилема мен флоэма арасындағы байланысты және байланысты қамтамасыз етеді, ал қанттар энергия көзі ретінде қауіпсіз түрде сақталады. Этилен деңгейі белсенді камбиальды зонасы бар өсімдіктерде жоғары және олар қазір де зерттелуде. Гиббереллин камбиальды жасушаның бөлінуін ынталандырады, сонымен қатар ксилема тіндерінің дифференциациясын реттейді, бұл флоэманың дифференциалдау жылдамдығына әсер етпейді. Дифференциация - бұл тіндерді арнайы түрге өзгертетін, өсімдіктің тіршілік формасын сақтауда маңызды рөл атқаратын маңызды процесс. Теректегі ағаштарда гиббереллиннің жоғары концентрациясы камбиальды жасушалардың бөлінуінің жоғарылауымен және камбиальды дің жасушаларында ауксиннің көбеюімен оң байланысты. Гиббереллин ксилеманың өсіндіден тамырға қарай таралатын сигнал арқылы кеңеюіне де жауапты. Цитокинин гормоны жасушалардың дифференциациясы бағытының орнына жасушаның бөліну жылдамдығын реттейтіні белгілі. Зерттеу көрсеткендей, мутанттардың сабағында және тамырда өсуінің төмендеуі анықталды, бірақ тамыр шоғырларының екінші реттік қан тамырлары цитокининмен емделмеген.[дәйексөз қажет ]
Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ Этчелс, Дж. Питер; Мишра, Лакси С .; Кумар, Манодж; Кэмпбелл, Лиам; Тернер, Саймон Р. (сәуір 2015). «Ағаштардағы ағаштардың түзілуі PXY-реттелетін жасушалар бөлігін манипуляциялау арқылы көбейеді». Қазіргі биология. 25 (8): 1050–1055. дои:10.1016 / j.cub.2015.02.023. PMC 4406943. PMID 25866390.
Сыртқы сілтемелер
- Суреттері Тамырлы камбий
- Толық сипаттама - Джеймс Д. Маузет
- Шолу; Risopatron, JPM; Күн, YQ; Джонс, BJ (2010). «Тамырлы камбий: Жасуша құрылымын молекулалық бақылау». Протоплазма. 247 (3–4): 145–161. дои:10.1007 / s00709-010-0211-з. PMID 20978810. S2CID 21775569.