Полисомды профильдеу - Polysome profiling

Полисомды профильдеу ішіндегі техника молекулалық биология ассоциациясын зерттеу үшін қолданылады мРНҚ бірге рибосомалар. Бұл техниканың өзгеше екенін ескеру маңызды рибосома профилі. Екі техника да қарастырылды[1] және екеуі де талдау кезінде қолданылады аудармашы, бірақ олар шығаратын мәліметтер спецификаның әр түрлі деңгейлерінде болады. Сарапшылар қолданған кезде, техника керемет қайталанатын болып табылады: бірінші кескіндегі 3 профиль 3 түрлі тәжірибеден алынған.[2]
Процедура
Процедура a жасаудан басталады жасуша лизаты қызығушылық ұяшықтарының. Бұл лизат құрамында полисомалар, моносомалар (құрамында орналасқан бір рибосомадан тұрады мРНҚ ), кішкентай (40S дюйм) эукариоттар ) және үлкен (эукариоттарда 60S) рибосомалық суббірліктер, «бос» мРНҚ және көптеген басқа еритін жасушалық компоненттер.
Процедура үздіксіз жасау арқылы жалғасады сахароза а-дағы үздіксіз айнымалы тығыздық градиенті центрифуга түтік. Қолданылған концентрацияда (мысалда 15-45%) сахароза рибосомалар мен мРНҚ ассоциациясын бұзбайды. Градиенттің 15% бөлігі түтіктің жоғарғы жағында, ал 45% бөлігі төменгі жағында орналасқан, өйткені олар әр түрлі тығыздық.
Белгілі бір мөлшер (өлшенгендей) оптикалық тығыздық ) содан кейін лизат түтіктегі градиенттің үстіне ақырын қабаттасады. Лизат, құрамында көп мөлшерде еритін материал болса да, 15% сахарозадан әлдеқайда аз тығыз, сондықтан оны ақырын жасаған жағдайда түтіктің жоғарғы жағында бөлек қабат ретінде ұстауға болады.
Лизаттың компоненттерін бөлу үшін препарат центрифугалауға ұшырайды. Бұл жылдамдатады лизаттың құрамдас бөліктері ауырлық күші және осылайша оларды жеке компоненттердің қаншалықты «үлкен» екендігіне негізделген градиент арқылы қозғалтады. Кішкентай (40S) суббірліктер градиентке үлкен (60S) суббірліктерге қарағанда аз жүреді. МРНҚ-дағы 80S рибсомалары әрі қарай жүреді (mRNA мөлшерінің жүріп өткен қашықтыққа қосқан үлесі айтарлықтай емес екенін ескеріңіз). 2 рибосомадан тұратын полисомалар әрі қарай жүреді, 3 рибомадан тұратын полисомалар қозғалмай әрі қарай жүреді. Компоненттердің «өлшемін» S, the белгілейді сведберг бірлік. Бірі S = 10 екенін ескеріңіз−13 секунд, және бұл «үлкен» ұғымы шындығында тым жеңілдету болып табылады.

Центрифугалаудан кейін пробирканың мазмұны жоғарыдан фракциялар ретінде жиналады (кішірек, баяу қозғалады) төменге қарай (үлкен, жылдам жүреді) және фракциялардың оптикалық тығыздығы анықталады. Алынған алғашқы фракцияларда салыстырмалы түрде аз молекулалардың көп мөлшері бар, мысалы, тРНҚ, жеке белоктар және т.б.
Қолданбалар
Бұл техниканы ұяшықтардағы жалпы аударма дәрежесін зерттеу үшін қолдануға болады (мысалдар үшін)[3][4][5]), бірақ оны жекелеген белоктар мен олардың мРНҚ-ын зерттеу үшін нақтырақ қолдануға болады. Суреттің төменгі бөлігінде көрсетілген мысал ретінде, кіші суббірліктің бір бөлігін құрайтын ақуызды алдымен 40S фракциясында анықтауға болады, содан кейін 60S фракциясынан жоғалады (бұл градиенттердегі бөліністер абсолютті емес), содан кейін қайтадан пайда болады 80S және полисомалық фракцияларда. Бұл жасушада кіші суббірлікке кірмейтін ақуыздың ең аз мөлшері бар екенін көрсетеді. Керісінше, иммуноблот фигурасының жоғарғы қатарында еритін фракцияларда еритін ақуыз пайда болады және рибосомалар мен полисомалармен байланысты. Белгілі бір а шаперон ақуызы, бұл (қысқаша) жаңа туылған нәрестені бүктеуге көмектеседі пептид өйткені ол рибосомадан шығарылып жатыр. Басқа жұмыс сияқты
көрсетілген қағазда шаперонның рибосомамен тікелей байланысы бар.[2]
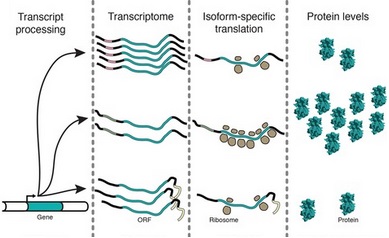
Бұл әдістемені белгілі бір мРНҚ-ның аударылу дәрежесін зерттеу үшін де қолдануға болады[6] Бұл тәжірибелерде мРНҚ-ның 5 'және 3' тізбектері олардың өндірілген мРНҚ мөлшеріне және мРНҚ-ның қаншалықты жақсы аударылғанына әсер етуі зерттелді. Көрсетілгендей, барлық mRNA изоформалары бірдей тиімділікпен аударылмайды Сөйтсе де олардың кодтау реттілігі бірдей.[6]
Пайдаланылған әдебиеттер
- ^ Пичирилло, Калифорния; т.б. (2014). «Иммундық жауаптардың трансляциялық бақылауы: транскрипциядан трансляторомға дейін». Табиғат иммунологиясы. 15 (6): 503–511. дои:10.1038 / ni.2891. PMID 24840981.
- ^ а б Ханебут, MA; т.б. (2016). «Hsp70 Ssb мультивалентті контактілері оның рибосомалардағы сәулеленуіне және туындайтын тізбектің өзара әрекеттесуіне ықпал етеді». Табиғат байланысы. 7: 13695. Бибкод:2016NatCo ... 713695H. дои:10.1038 / ncomms13695. PMC 5150220. PMID 27917864.
- ^ Лин, Дж .; т.б. (2010). «Антидепрессант сертралин рапамицин сигнализациясының сүтқоректілердің нысанын қысқарту арқылы трансляцияның басталуын тежейді». Онкологиялық зерттеулер. 70 (8): 3199–3208. дои:10.1158 / 0008-5472.CAN-09-4072. PMID 20354178.
- ^ Коудерт, Л; т.б. (2014). «Полисомдық профильдеу арқылы стресс жағдайындағы аударманың басталуын талдау». Көрнекі тәжірибелер журналы (87). дои:10.3791/51164. PMC 4193336. PMID 24893838.
- ^ Молон, М; т.б. (2016). «Saccharomyces cerevisiae ашытқысының ұзақ өмір сүруін анықтайтын фактор ретінде метаболизм жылдамдығы». Жасы (Дордрехт, Нидерланды). 38 (1): 11. дои:10.1007 / s11357-015-9868-8. PMC 5005888. PMID 26783001.
- ^ а б Еден, ЕН; Doudna, JA (2016). «Адам жасушаларында транскриптивті изоформалар арқылы реттелетін ақуыз синтезі». eLife. 5. дои:10.7554 / eLife.10921. PMC 4764583. PMID 26735365.