Фомоксантон A - Phomoxanthone A
 | |
| Атаулар | |
|---|---|
| IUPAC атауы [(3R, 4R, 4aR) -5 - [(5R, 6R, 10aR) -5-Ацетилокси-10а- (ацетилоксиметил) -1,9-дигидрокси-6-метил-8-оксо-6,7-дигидро- 5Н-ксантен-4-ыл] -4-ацетилокси-8,9-дигидрокси-3-метил-1-оксо-3,4-дигидро-2Н-ксантен-4а-ыл] метил ацетат | |
| Басқа атаулар PXA | |
| Идентификаторлар | |
3D моделі (JSmol ) | |
| Чеби | |
| ЧЕМБЛ | |
| ChemSpider | |
PubChem CID | |
| |
| |
| Қасиеттері | |
| C38H38O16 | |
| Молярлық масса | 750,70 г / моль |
| Сыртқы түрі | сары қатты |
| Тығыздығы | ~ 1,53 г / см3 |
| ерімейді | |
| Ерігіштік жылы DMSO | жақсы, бірақ тұрақсыз[1] |
| Ерігіштік жылы EtOH | орташа[1] |
Өзгеше белгіленбеген жағдайларды қоспағанда, олар үшін материалдар үшін деректер келтірілген стандартты күй (25 ° C [77 ° F], 100 кПа). | |
| Infobox сілтемелері | |
The микотоксин фомоксантон A, немесе PXA қысқаша айтқанда, уытты болып табылады табиғи өнім әсер етеді митохондрия. Бұл ең улы және табиғи түрде ең жақсы зерттелген фомоксантондар. Жақында PXA жылдам, канондық емес индукциялауға ұшырады митохондриялық бөліну тудыруы арқылы митохондриялық матрица фрагментке дейін сыртқы митохондриялық мембрана өзгеріссіз қалуы мүмкін. Бұл процесс митохондриялық бөлінуден және тәуелсіз екендігі көрсетілген біріктіру реттеушілер DRP1 және OPA1.[1]
Қасиеттері мен құрылымы

Фомоксантондар саңырауқұлақтың атымен аталады Фомопсис, олар алдымен оқшауланған және олардан кейін ксантоноид құрылым, демек олардың қосылысқа ұқсас құрылымдары бар ксантон (суретте сол жақта). Химиялық тұрғыдан алғанда, фомоксантондар екі тетрагидроксантонның димерлері, яғни олар төрт ксантоноидтардың екі суббірлігінен тұрады. гидрокси топтары әрқайсысы. Фомоксантондардың екі суббірлігі ковалентті бір-бірімен байланысты. PXA өзі гомодимер болып табылады, яғни ол екі бірдей суббірліктен тұрады. Бұл екі бөлімшелер де диацетилденген тетрагидроксантондар, сондықтан олардың екі гидрокси тобы ауыстырылды ацетил топтары. Екі димерлі суббірліктің арасындағы байланыс позициясы PXA мен оның аз уытты изомерлері арасындағы жалғыз құрылымдық айырмашылық болып табылады фоноксантон B (PXB) және дикерандрол C: PXA-да екі ксантоноидты мономерлер C-4,4 'позициясында симметриялы түрде байланысады, ал PXB-да олар C-2,4', және дикерандрол C-де олар C-2-де симметриялы байланысады , 2 '. Әйтпесе, бұл үш қосылыс құрылымдық жағынан бірдей.[2][3] Фомоксантондар құрылымдық жағынан тығыз байланысты секалон қышқылдары, димерлі тетрагидроксантон микотоксиндерінің тағы бір класы, олар бірнеше қасиеттерге ие. Фомоксантондар да, секалон қышқылдары да еріген кезде тұрақсыз полярлы еріткіштер сияқты DMSO, бірге ковалентті байланыс 2,2′-, 2,4′- және 4,4′-байланыс арасында ауысатын екі мономердің арасында.[4] Екі PXA және PXB фомоксантоны баяу жүре алады изомерия бір-біріне, сондай-ақ, улы емес дикерандрол С-ға түсіп, нәтижесінде полярлы еріткіште еріген кезде PXA белсенділігі жоғалады.[1]
Пайда болу
Қалай табиғи өнімдер, PXA және басқа фомоксантондар пайда болады екінші метаболиттер аттас тектегі саңырауқұлақтарда Фомопсис, әсіресе түрде Phomopsis longicolla.[2][3] Бұл саңырауқұлақ ан эндофит мангров зауытының Sonneratia caseolaris.[5][3] Сонымен қатар, ол басқа өсімдіктерде қоздырғыш ретінде анықталған, мысалы соя деп аталатын ауруды тудыратын өсімдік Фомопсис тұқымның ыдырауы (PSD).[6][7]
Дайындық
PXA және PXB екеуі де 2001 жылы ашылды және оларды оқшаулау арқылы дайындау Фомопсис саңырауқұлақ дақылдары тиісті басылымда сипатталған.[2] Қысқаша, а MeOH сығындысы Фомопсис мәдениет араласады H2O және жуылады гексан. Содан кейін сулы фазаны кептіріп, қалдықты ерітеді EtOAc, H-мен жуылады2О, концентрацияланған және бірнеше рет тазартылған өлшемді-алып тастау хроматографиясы. Алынған PXA мен PXB қоспасы бөлінеді HPLC. Бастапқы экстракция MeOH орнына EtOAc көмегімен жүзеге асырылатын және кептіру қадамы өткізіп жіберетін өзгертілген әдіс 2013 жылы сипатталған.[3]
Қолданады
Фомоксантон А алғаш рет скринингте анықталды безгекке қарсы қосылыстар.[2] Бұл антибиотиктің а-ға қарсы белсенділігін көрсетті есірткіге төзімді негізгі қоздырғышының штаммы безгек, қарапайым паразит Plasmodium falciparum. Сол зерттеу PXA антибиотиктік белсенділігі туралы хабарлады Туберкулез микобактериясы және жануарлардың жасушаларының үш сызығына қарсы, олардың екеуі адамның рак клеткаларынан алынған.[2] Бұл нәтижелер PXA-ның әр түрлі организмдерге қарсы антибиотикалық белсенділігі бар екенін көрсетіп қана қоймай, сонымен қатар PXA-ны әлеует ретінде зерттейтін зерттеулерге де себеп болды. антибиотик немесе қатерлі ісікке қарсы препарат. Кейінгі зерттеу сонымен қатар PXA-ға қарсы антибиотиктердің балдырларға қарсы белсенділігі туралы хабарлады Chlorella fusca, саңырауқұлақтар Устилаго шегірткесі және бактерия Bacillus megaterium.[8] Бұл белсенділіктің кең ауқымы оны емдеуде қолдануға болатын арнайы антибиотик ретінде жарамсыз етті жұқпалы аурулар дегенмен, оны қатерлі ісікке қарсы дәрі ретінде пайдалануға болады деген үміт қалды. Адамның қатерлі ісік жасушалары мен қатерлі ісікке жатпайтын жасушаларында жүргізілген зерттеудің алғашқы нәтижелері PXA екіншісіне қарағанда біріншісіне улылырақ болуы мүмкін деген болжам жасады, дегенмен in vivo зерттеулерінің нәтижелері әлі ұсынылмаған.[3][9]
Потенциалды медициналық қолданудан басқа, соңғы зерттеулер PXA митохондриялық мембрананың динамикасын, әсіресе канондық емес митохондриялық бөлінуді және митохондриялық матрицаны қайта құруды зерттеу құралы ретінде қолдануы мүмкін екенін көрсетеді.[1]
Биологиялық белсенділік

PXA бактериялар, протозойлар, саңырауқұлақтар, өсімдіктер мен жануарлар жасушалары сияқты әр түрлі организмдерге қарсы антибиотиктік белсенділікке ие болғандықтан, адамның қатерлі ісігі жасушаларын жасушалық ерекшелікке әсер етуі керек. Жақында жүргізілген зерттеу көрсеткендей, PXA митохондрияға олардың биохимиялық функцияларын да, мембраналық архитектурасын да бұзу арқылы тікелей әсер етеді.[1] Митохондриялар жасушалық болып табылады органоидтар барлығында бар эукариоттар. Теориясына сәйкес симбиогенез, олар бактериялардан алынған және олармен көптеген сипаттамаларды, соның ішінде олардың мембраналық құрамының бірнеше қасиеттерін бөліседі.[10][11]
Митохондрияның негізгі функцияларының бірі - жасушалық энергия валютасын өндіру ATP процесі арқылы тотығу фосфорлануы (OxPhos). OxPhos митохондрияға байланысты мембраналық потенциал арқылы жасалады электронды тасымалдау тізбегі Тұтыну арқылы (ETC) оттегі. PXA митохондрияның барлық осы функцияларына кедергі келтіретіні көрсетілген: бұл ATP синтезін төмендетіп, митохондрияны деполяризациялап қана қоймайды, сонымен қатар ETC және жасушалық оттегі шығынын тежейді. Бұл оны ерекшелендіреді агенттер сияқты протонофорлар. Бұлар ATP синтезін төмендетіп, митохондрияны деполяризацияласа да, мембрана потенциалын қалпына келтіру мақсатында ETC белсенділігінің жоғарылауына байланысты тыныс алуды бір уақытта күшейтеді.[1]
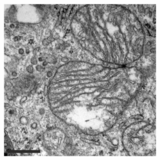
Митохондриялардың бұл тежелуінен басқа, PXA олардың мембраналық архитектурасын да бұзады. Көптеген жасуша типтерінде митохондриялар қалыпты түрде тепе-тең митохондриялық бөлінудің тұрақты процесіне ұшырайтын күрделі түтікшелі тор құрайды. митохондриялық біріктіру. PXA немесе көптеген басқа митохондриялық стрессорлармен емдеу, мысалы протонофоралар, митохондриялық фрагментацияға әкелетін шамадан тыс бөлінуді тудырады. PXA жағдайында бұл фрагментация процесі протонофоралар сияқты басқа агенттер тудырған канондық фрагментациядан бірнеше тәсілмен ерекшеленетіндігі көрсетілді: біріншіден, бұл жылдамырақ, нәтижесінде шамамен бір минут ішінде толық бөлшектену пайда болды Канондық фрагментация үшін 30-60 минут; екіншіден, ол митохондриялық бөлінуге және DRP1 және OPA1 біріктіру реттегіштеріне тәуелсіз; үшіншіден, PXA екеуінің де бөлшектенуіне әкеледі сыртқы митохондриялық мембрана (OMM) және митохондриялық матрица жабайы түрі бұл DRP1 жетіспейтін жасушаларда матрицаның эксклюзивті фрагментациясын тудырады.[1] Бұл соңғы ерекшелігі ерекше, өйткені матрицалық бөлінудің белсенді механизмі жоғары эукариоттарда белгілі емес.[12] Митохондриялық ультрақұрылымды зерттеу кезінде PXA пайда болатындығы анықталды кристалар митохондриялық матрицаның бұзылуы және толық бұрмалануы. Мүмкін, осы әсер арқылы PXA бағдарламаланған жасушалық өлімді келесі түрінде тудырады апоптоз.[1]
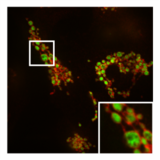
- PXA-ның митохондриялық морфологияға және ультрақұрылымға әсері
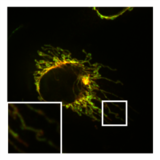
А-дағы митохондрияның қалыпты, түтікшелі морфологиясы ХеЛа бөліну медиаторы болатын жасуша DRP1 болды нокаут. Қосымша кескін митохондриялық матрица (жасыл) және OMM (қызыл).


Әдебиеттер тізімі
- ^ а б c г. e f ж сағ мен j Бёлер, Филип; Штулдрайер, Фабиан; Ананд, Ручика; Кондади, Арун Кумар; Шлютерманн, Дэвид; Берлет, Никлас; Дейтерсен, Яна; Уоллот-Хиеке, Нора; Ву, Вэнсянь; Фрэнк, Мариан; Ниманн, Гендрик; Весбер, Элизабет; Барбиан, Андреас; Люйтен, Томас; Парис, Ян Б; Вайдткамп-Питерс, Стефани; Борчардт, Андреа; Рейхерт, Андреас С; Пенья-Бланко, Аида; Гарсия-Саез, Ана Дж; Ицканов, Сэмюэль; Ван Дер Блик, Александр М; Прокш, Петр; Весселборг, Себастьян; Шторк, Бьорн (2018). «Микотоксиндік фомоксантон ішкі митохондриялық мембрананың формасы мен қызметін бұзады». Жасушалардың өлімі және ауруы. 9 (3): 286. дои:10.1038 / s41419-018-0312-8. PMC 5833434. PMID 29459714.
- ^ а б c г. e Исака, Масахико; Джатурапат, Амонлая; Руксерие, Камолчанок; Данвисетканьяна, Каннават; Тантичароен, Моракот; Thebtaranonth, Йодхатай (2001). «Фомоксантондар А және В, Эндофиттік саңырауқұлақтардан шыққан роман Ксантонның димерлері Фомопсис Түрлер «. Табиғи өнімдер журналы. 64 (8): 1015–8. дои:10.1021 / np010006h. PMID 11520217.
- ^ а б c г. e Рёнсберг, Дэвид; Деббаб, Абдессамад; Манди, Аттила; Васильева, Вера; Бёлер, Филип; Шторк, Бьорн; Энгельке, Лаура; Гамахер, Александра; Савадого, Ричард; Дидерих, Марк; Рэй, Виктор; Лин, Венхан; Кассак, Матиас У; Джаниак, Кристоф; Шеу, Стефани; Весселборг, Себастьян; Куртан, Тибор; Али, Амал Н; Proksch, Peter (2013). «Эндопитикалық саңырауқұлақтардан алынған про-апоптотикалық және иммуностимуляторлы тетрагидроксантонның өлшемдері Phomopsis longicolla". Органикалық химия журналы. 78 (24): 12409–25. дои:10.1021 / jo402066b. PMID 24295452.
- ^ Цинь, Тянь; Ивата, Такаюки; Төлем, Таня Т; Бутлер, Джон А; Porco, Джон А (2015). «Әр түрлі байланысы бар диметриялық тетрагидроксантондардың синтезі:» пішінді өзгерту «қасиеттерін зерттеу». Американдық химия қоғамының журналы. 137 (48): 15225–33. дои:10.1021 / jacs.5b09825. PMC 4863954. PMID 26544765.
- ^ Xing, X.K; Чен, Дж; Xu, MJ; Лин, ВХ; Guo, S.X (2011). «Қытайдың оңтүстік жағалауында орналасқан Sonneratia (Sonneratiaceae) мангро өсімдіктерімен байланысты саңырауқұлақ эндофиттері». Орман патологиясы. 41 (4): 334. дои:10.1111 / j.1439-0329.2010.00683.x.
- ^ Хоббс, Томас В; Шмиттеннер, А.Ф; Кутер, Джеффри А (1985). «Соядан алынған жаңа фомопсис түрлері». Микология. 77 (4): 535. дои:10.2307/3793352. JSTOR 3793352.
- ^ Ли, Шуциан; Дарвиш, Омар; Альхаруф, Надим В; Мусунгу, Брайан; Мэттьюс, Бенджамин Ф (2017). «Phomopsis longicolla геномының реттілігін талдау: соя бұршағында Phomopsis тұқымдарының ыдырауын тудыратын саңырауқұлақ қоздырғышы». BMC Genomics. 18. дои:10.1186 / s12864-017-4075-x. PMID 28870170.
- ^ Эльзассер, Бригитта; Крохн, Карстен; Флёрке, Ульрих; Тамыр, Наталья; Ост, Ганс-Юрген; Дрегер, Зигфрид; Шульц, Барбара; Антус, Шандор; Куртан, Тибор (2005). «Фоноксантон А-ның рентген құрылымын анықтау, абсолютті конфигурациясы және биологиялық белсенділігі». Еуропалық органикалық химия журналы. 2005 (21): 4563. дои:10.1002 / ejoc.200500265.
- ^ Фрэнк, М; Ниманн, Н; Бёлер, П; Лейлек, B; Весселборг, С; Лин, В; Proksch, P (2015). «Фомоксантон А - мангроу ормандарынан қатерлі ісікке қарсы терапияға дейін». Қазіргі дәрілік химия. 22 (30): 3523–32. дои:10.2174/0929867322666150716115300. PMID 26179997.
- ^ Мартин, Уильям Ф; Гарг, Срирам; Зиморский, Верена (2015). «Эукариоттың шығу тегі туралы эндосимбиотикалық теориялар». Корольдік қоғамның философиялық операциялары В: Биологиялық ғылымдар. 370 (1678): 20140330. дои:10.1098 / rstb.2014.0330. PMC 4571569. PMID 26323761.
- ^ Милейковская, Е; Dowhan, W (2009). «Прокариоттар мен эукариоттардағы кардиолипинді мембраналық домендер». Biochimica et Biofhysica Acta. 1788 (10): 2084–2091. дои:10.1016 / j.bbamem.2009.04.003. PMC 2757463. PMID 19371718.
- ^ Ван Дер Блик, А.М; Шен, Q; Каваджири, С (2013). «Митохондриялық бөліну және бірігу механизмдері». Биологиядағы суық көктем айлағының болашағы. 5 (6): a011072. дои:10.1101 / cshperspect.a011072. PMC 3660830. PMID 23732471.
Сыртқы сілтемелер
 Қатысты медиа Фомоксантон Wikimedia Commons сайтында
Қатысты медиа Фомоксантон Wikimedia Commons сайтында