Жынды1 - Mad1 - Wikipedia
| Жынды1 | |||||||
|---|---|---|---|---|---|---|---|
 Хрусталь құрылымы, Mad1-Mad2 кешенінің тетрамері, сары және қызыл = Mad1 мономерлері, ақшыл жасыл = Mad2 мономерлері | |||||||
| Идентификаторлар | |||||||
| Организм | |||||||
| Таңба | MAD1 | ||||||
| Энтрез | 852794 | ||||||
| PDB | 1GO4 | ||||||
| RefSeq (mRNA) | NM_001180951.3 | ||||||
| RefSeq (прот) | NP_011429.3 | ||||||
| UniProt | P40957 | ||||||
| Басқа деректер | |||||||
| Хромосома | VII: 0,35 - 0,35 Мб | ||||||
| |||||||
Жынды1 маңызды емес ақуыз қайда ашытқы функциясы бар шпиндельді жинауды бақылау пункті (SAC).[1]Бұл бақылау нүктесі шпиндель микротүтікшелеріне хромосомалардың қосылуын бақылайды және шпиндель салынғанша жасушалардың анафазаның басталуына жол бермейді. Mad атауы микро түтікшелер деполимеризациясы кезінде мутантты жасушалардың митотикалық тоқтату тапшылығы (MAD) болатындығын бақылауды білдіреді. Mad1 анафаза тежегішін қабылдайды Жынды2 бекітілмеген кинетохорларға және Mad2- үшін өте қажетCD20 күрделі формация in vivo бірақ жоқ in vitro. In vivo, Mad1 Mad2-Cdc20 кешенінің бәсекеге қабілетті ингибиторы ретінде әрекет етеді.[2] Mad1 Mps1 арқылы фосфорланады, содан кейін басқа әрекеттермен бірге митотикалық бақылау-өткізу кешенінің (MCC) пайда болуына әкеледі. Осылайша ол анафазаға ықпал ететін комплекс / циклосоманың (АПС / С) белсенділігін тежейді. Mad1 гомологтары эукариоттарда ашытқыдан сүтқоректілерге дейін сақталады.
Кіріспе
90-шы жылдардың басында ашытқы гендері анықталды, олар мутация нәтижесінде микротүтікшені бөлшектеуге жауап ретінде митозды тоқтату ақауларына әкелді (митотоксикалық жетіспейтін гендер - MAD гендері). Бұл жасушаларда микротүтікшелі полимеризация ингибиторлары болған кезде митоздық тоқтату байқалмады, сондықтан жасушалардың бөлінуін кешіктіре алмады.[1] Анықталған гендер құрамына кірді MAD1, MAD2 және MAD3 гендер. Олар бәрінде сақталған эукариоттар және белсенді болатын жолға қатысады прометафаза қарындасының мерзімінен бұрын бөлінуіне жол бермеу хроматидтер және деп аталатындарды құрайды шпиндельді жинауды бақылау пункті (SAC). Бұл бақылау пункті хромосоманың митотикалық шпиндельге қосылу күйін бақылайды және ингибирлейді метафаза дейін анафаза активациясының алдын-алу арқылы өту анафазаға ықпал ететін кешен / циклосома (APC / C) және сол арқылы деградация туралы жасушалық цикл реттеушілер.[3] Mad1 бұл жолда бекітілмеген кинетохорларда жинақталған және осы машинадағы бекітілмеген кинетохорлар үшін сенсор қызметін атқарады.
Функция
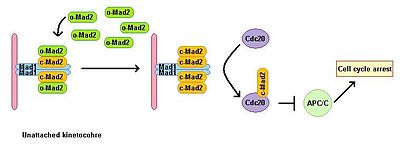
Эукариотты жасушалар микротүтікшелі полимерлену ингибиторларының қатысуымен митоздық тоқтауды көрсетеді. Шпиндельді жинауды бақылау нүктесі шпиндельдің күйін бақылайды және метафаза-анафазаның ауысуын барлық кинетохорлардың митотикалық шпиндельге дұрыс биполярлы бекітілуіне байланыстырады. Шпиндельді жинауды бақылау нүктесі анафазаның басталуы мен митоздан шығуына алып келетін төменгі эффекторлардың деградациясын болдырмау арқылы анафазаны алға жылжытатын кешеннің белсенділігін тежейді. Mad1 сарқылуы жоғалтуға әкеледі МАК функциясы. Mad1 негізінен бекітілмеген кинетохорларда локализацияланады және бірыңғай бекітілмеген кинетохор жағдайында митоздық қамауға әкеледі. Mad1 маңызды SAC компонентін алады Жынды2 бекітілмеген кинетохорларға және митоздық тоқтату сигналының күшеюіне әкеледі. Оның белсенді емес ашық конформациясында бос цитоплазмалық Mad2 бассейні бар, ол o-MAD2 деп аталады. Mad1-мен байланысқан кезде, Mad2 белсенді қабылдайды конформация жабық деп аталады (с-Mad2) және екі Mad1 және екі с-Mad2 бірлігінің гетеротетремерін құрайды. Mad1 – c-Mad2 гетеротетрамері өте тұрақты және а ретінде жұмыс істейді каталитикалық рецептор ақысыз цитоплазмалық o-Mad2 үшін. Еркін o-Mad2 осы рецептормен байланысады және оның конформациясын белсенді жабық түрге өзгертеді. Бұл екінші c-MAD2 көшіріледі CD20 әлі белгісіз механизмі бар және формалары Cdc20 – c-Mad2. Бұл кешен митотикалық бақылау-өткізу кешенінің (МКК) маңызды компоненті болып табылады. MCC байланыстырады және тежейді APC / С, сондықтан митоз арқылы прогрессияны тоқтатады.[3][4]
Реттеу
Ағысқа қарсы екі бақылау бекеті бар киназалар арқылы Mad1 функциясын реттеуге қатысады фосфорлану.[5] Mps1 Mad1-ді де фосфорлайды in vitro және in vivo және Mad1 және Mad2 локализациясын реттейді деп ойлайды кинетохорлар және олардың өзара әрекеттесу динамикасы. BUB1 Mad1-ді кинетохорға қабылдайтын және кинетохор бекітілмеген болса, оны белсендіретін басқа киназа.[3]Егер кинетрохор шпиндельге бекітілген болса, онда SAC ингибиторы p31құйрықты жұлдыз Mad2-нің Mad2-дің конформациялық қайта құрылымын тежейді және Mad2-дің Cdc20-мен байланысуына жол бермейді.[3]
Құрылымдық ерекшеліктері мен механизмі

Биохимиялық әдістермен Mad1 90кД, 718 қалдықты кодтайды деп болжанған,[6] ширатылған катушка ақуыз тән таяқша пішінімен[1]1995 жылы. Көп ұзамай кристалды құрылымдар пайда болды. Содан кейін 2002 жылы тетрамер түзетін адам Mad2-мен бірге адамның Mad1 кристалдық құрылымы жарық көрді. Эксперименттік шектеулерге байланысты құрылым тек Mad1 қалдықтарын 484 - 584 көрсетеді. Ұзартылған Mad1 мономерлері N терминалы альфа спиральдарының қатысуымен параллель ширатылған катушкамен тығыз орналасқан. Mad1 тізбектері ширатылған катушкадан Mad2 лигандарына қарай Mad2-мен екі қосалқы комплекс түзеді. Альфа-спиральдар 1 мен 2 арасындағы сегмент Mad2 байланыстыру доменін қамтиды. Бұл байланыстырушы доменнің бірінші бөлігі икемді және асимметриялық кешенді тудыратын әр түрлі конформацияларды қабылдайды. Термодинамикалық зерттеулерді қолдана отырып, өз жұмыстарында Сирони және басқалар.[2]Mad1 функциялары Mad2- жылдамдығын бәсеңдету сияқты екенін көрсетедіCD20 күрделі қалыптасу, сондықтан бәсекеге қабілетті ингибитор рөлін атқарады in vivo. Сонымен қатар, авторлар Mad1-Mad2 байланыстыратын алаңдар құрылымның ішіне көміліп, байланыстырушы орындарды Cdc20 байланыстыруға қол жетімсіз етіп шығарады. Mad1-Mad2 байланысы әдеттен тыс, өйткені Mad2 C-терминалы Mad1 үстінен қатпарланады. Авторлар, демек, мазаланбаған Mad1-Mad2 кешені конформациялық өзгеру механизмін, әлі де болса жете түсінілмеген, романды қажет ететін Mad2 шығармайды деп тұжырымдайды.[2]
Қатерлі ісік
Мейоз кезінде хромосома санындағы сәйкессіздіктер (анеуплоидиялар) Даун синдромы сияқты адамның ауруына жауап береді және рак клеткаларында жиі пайда болады. SAC маңызды функциясы SAC мутациясы және әсіресе SAC инактивациясы туморигенезге себеп болуы немесе ең болмағанда тумигенезді жеңілдетуі мүмкін деген гипотезаны тудырады.[3] Бұл идеяға қарсы рак клеткалары МАК компоненттері болмаған кезде апоптозға ұшырайтындығы көрсетілген.[7] Бұл модельде, басқа модельден айырмашылығы, SAC инактивациясы тез бөлінетін рак клеткаларын жоюдың әлеуетті әдісіне айналады. Mad1p, SAC, апоптоз және қатерлі ісік арасындағы молекулалық байланыстар әлі де толық зерттелмеген.[3]
Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ а б c Hardwick KG, Murray AW (1995). «Mad1p, ашытқы жасушасында шпиндельді жинауды бақылау пунктінің фосфопротеинді компоненті». Жасуша биологиясының журналы. 131 (3): 709–720. дои:10.1083 / jcb.131.3.709. PMC 2120625. PMID 7593191.
- ^ а б c Sironi L, Mapelli M, Knapp S, De Antoni A, Jeang KT, Musacchio A (2002). «Mad1-Mad2 тетрамерикалық ядролық кешенінің кристалдық құрылымы: шпиндельді бақылау пункті үшін» қауіпсіздік белдігін «байланыстыру механизмінің салдары». EMBO журналы. 21 (10): 2496–2506. дои:10.1093 / emboj / 21.10.2496. PMC 126000. PMID 12006501.
- ^ а б c г. e f Musacchio A, Лосось ED (мамыр 2007). «Кеңістіктегі және уақыттағы шпиндельді-жинау пункті». Нат. Аян Мол. Жасуша Биол. 8 (5): 379–93. дои:10.1038 / nrm2163. PMID 17426725. S2CID 205494124.
- ^ Ю Х (сәуір 2006). «Митозды шпиндельді бақылау нүктесінде Mad2 құрылымдық активациясы: екі күйлі Mad2 моделі мен Mad2 шаблонының моделі». Дж. Жасуша Биол. 173 (2): 153–157. дои:10.1083 / jcb.200601172. PMC 2063805. PMID 16636141.
- ^ Bharadwaj R, Yu H (2000). «Шпиндельді бақылау нүктесі, анеуплоидия және қатерлі ісік». Онкоген. 23 (11): 2016–27. дои:10.1038 / sj.onc.1207374. PMID 15021889.
- ^ Чен Р.Х., Шевченко А, Манн М, Мюррей А.В. (1998). «Spindle Checkpoint Protein Xmad1 Xmad2-ді бекітілмеген кинетохорларға қабылдайды». Жасуша биологиясының журналы. 143 (2): 283–295. дои:10.1083 / jcb.143.2.283. PMC 2132829. PMID 9786942.
- ^ Kops GJ, Foltz DR, Кливленд DW (маусым 2004). «Митозды бақылау нүктесін тежеу арқылы хромосоманың көп мөлшерде жоғалуы арқылы адамның қатерлі ісік жасушаларына өлім». Proc. Натл. Акад. Ғылыми. АҚШ. 101 (23): 8699–704. Бибкод:2004PNAS..101.8699K. дои:10.1073 / pnas.0401142101. PMC 423258. PMID 15159543.