Гомосинапстық икемділік - Homosynaptic plasticity - Wikipedia
Гомосинапстық икемділік бір түрі болып табылады синаптикалық икемділік.[1] Гомосинапстық икемділік кіріске тән, яғни синапстың беріктігінің өзгеруі синапстыққа дейінгі мақсатпен арнайы ынталандырылған синапстықтан кейінгі мақсаттарда ғана болады.[2] Сондықтан синапстыққа дейінгі ұяшықтан сигналдың таралуы локализацияланған.
Синаптикалық пластиканың тағы бір түрі, гетеросинапстық икемділік, кіріске тән емес және көптеген механизмдер бойынша гомосинапстық пластикадан ерекшеленеді.
Гомосинапстық икемділік арқылы синапсты күшейту ассоциативті болып табылады, өйткені ол пресинапстық және постсинапстық нейронның уақытында атылуына байланысты. Бұл ассоциативтілік постсинаптикалық нейронның өртену мүмкіндігін арттырады.[3] Бұл механизмдер оқытудың негізінде теорияланған қысқа мерзімді жады.[3]
Шолу
Хеббтің постулаты
Дональд Хебб синаптикалық байланыстардың күшеюі синапстыққа дейінгі терминал мен синаптикалық дендрит арасындағы үйлестірілген белсенділіктің арқасында пайда болды деген теорияны алға тартты. Хебб бойынша, бұл екі жасуша күшейе түседі, өйткені олардың сигналдары кеңістікте және / немесе уақытта бірге жүреді, оларды кездейсоқ әрекет деп те атайды. Бұл постулатты жиі қысқаша сипаттайды Бірге жанатын ұяшықтар, бір-бірімен сымдемек, кездейсоқ атудан нейрондары бар синапстар күшейтіледі, ал осы нейрондардағы басқа синапстар өзгеріссіз қалады.[3] Хеббтің постулаты ұзақ мерзімді ақпаратты сақтаудың синаптикалық икемділігі негізінде тұжырымдамалық негіз құрды.[1]
Кірістің ерекшелігі механизмдері
Икемділіктің өзгеруі көбінесе AMPA рецепторларын (AMPAR) синапстың постсинаптикалық мембранасына дәнекерлеу күшінің өзгеруімен енгізу немесе интерьеризациялау арқылы жүреді.[1] Ca2+ - бұл жасуша ішіндегі биологиялық өзгерістердің каскадын индукциялау арқылы осы AMPA рецепторларының тығыздығының өзгеруін тудыратын сигналдық ион. Индукциялау үшін ұзақ мерзімді потенциал (LTP), Ca2+ CAMKII және PKC активтендіреді, бұл фосфорлануға және AMPAR енгізілуіне әкеледі ұзақ мерзімді депрессия (LTD) Ca арқылы жүреді2+ депосфорилаттайтын және АМПАР-ның интерьеризациясын тудыратын протеин фосфатазаларын белсендіреді.[1]
Синаптикалық беріктікке енгізілімге тән өзгерістер жасау үшін, Са2+ сигнал белгілі дендриттік тікенектермен шектелуі керек. Ca-ның дендритикалық шектелуі2+ бірнеше механизмдер арқылы жүзеге асырылады. Жасушадан тыс Ca2+ омыртқаға NMDA рецепторлары (NMDAR) және кернеу қақпасы арқылы ене алады2+ арналар (VGCC). NMDAR де, VGCC де дендриттік омыртқаларға шоғырланған, аралық омыртқаға Ca2+ ағын. Ca-ның жасушалық дүкендері2+ эндоплазмалық тор мен митохондрияда омыртқада шектеулі сигнализация пайда болуы мүмкін, дегенмен кейбір зерттеулер бұған дәлел таба алмады.[4] Са тазарту2+ Са-мен байланысатын буферлік белоктармен бақыланады2+ және оны басқа тікенектерге жол бермеңіз. Ca-ның шектеулі диффузиясы2+ дендритикалық омыртқаның мойнында оны белгілі бір дендриттермен оқшаулауға көмектеседі.[4]
Кіріске арналған ұзақ мерзімді әлеуеттің тағы бір механизмі уақытша болып табылады. NMDAR деполяризацияны, магний блогын алып тастауды және глутамат активациясын, өз арналарын ашып, Са2+ ағын. LTP осылайша NMDA арналары глутаматты шығаратын және постсинаптикалық жасушаның деполяризациясын тудыратын белсенді синаптикалық кірістермен ашылатын жерлерде локализацияланған және жақын орналасқан белсенді емес синапстарға әсер етпейді.[1]
Ұзақ мерзімді өзгерістерді сақтау
LTP-ді тұрақтандыру және оны ұзақ уақытқа созу үшін бұл өзгерісті қолдайтын жаңа ақуыздар күшейтетін синапстағы ынталандыруға жауап ретінде синтезделеді. Қиындық туындайды, синтезделген протеиндерді қажетті синапстарға қажетті синапстарға қалай алу керек. Бұл мәселені шешудің екі жолы бар синаптикалық тегтеу және жергілікті ақуыз синтезі.
Синаптикалық тегтеу
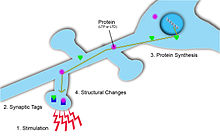
Синаптикалық тегтер синаптикалық икемділік пайда болған жерді белгілеңіз және бұл синаптикалық беріктік пен ұзақ мерзімді пластикалық өзгерістердің әлеуеті туралы ақпарат бере алады.[5] Тег уақытша болып табылады және оған Са ағынымен белсендірілген көптеген ақуыздар кіреді2+ постсинапстық жасушаға.[5] Сонымен қатар, синаптикалық өзгерістің типі мен шамасына байланысты, тегтеу үшін әр түрлі белоктар қолданылады. Мысалы, пластикалық өзгерістер ұзақ мерзімді депрессияға әкелгенде, кальциневрин қолданылады. Керісінше, икемділік ұзақ мерзімді күшейтуге әкелсе, CaMKII қолданылады.[5] Синаптикалық икемділік кіріске тән болуы үшін, синаптикалық әлеуеттің локализацияланған болуын қамтамасыз ету үшін, синаптикалық пост-синаптикалық мақсаттар үшін маңызды болып табылады.[5] Бұл тегтер кейінірек белок синтезін бастайды, бұл өз кезегінде осы активтендірілген нейрондарда синаптикалық пластиканың өзгеруіне әкеледі.[1]
Жергілікті ақуыз синтезі
Ақуыз синтезі дендриттер гомосинапстық икемділік үшін қажет. Постсинапстық жасушадағы деполяризация және нәтижесінде AMPA және NMDA рецепторларының активтенуі осы рецепторлардың эндоцитозын тудырады. Синапста беткі рецепторлардың санын ұстап тұру үшін жергілікті ақуыз синтезі қажет. Бұл жаңа белоктар гомосинаптикалық икемділіктің әсерінен болатын құрылымдық өзгерістерді тұрақтандыруға көмектеседі.[6] Дендриттерде осы ақуыздарды өндіре алатын рибосомалар бар. Сонымен қатар, дендриттерде РНҚ түйіршіктері бар, бұл жаңадан жасалған ақуыздардың болуын көрсетеді. LTP-ні синаптический мақсатты нейронның сомасынан бөлінген дендриттерден шақыруға болады. Керісінше, LTP осы дендриттерде Endomyacin сияқты ақуыз синтезінің блокаторларымен бұғатталуы мүмкін, бұл әрі қарай жергілікті ақуыз синтезделетін орынды көрсетеді. Бұл дәлел жергілікті ақуыз синтезінің L-LTP тұрақтандырылуы және сақталуы үшін қажет екенін көрсетеді.[1]
Әдебиеттер тізімі
- ^ а б c г. e f ж Первс, Д., Августин, Дж., Фицпатрик, Д., Холл, В.С., ЛаМантиа, А.С., Уайт, Л.Э. (2012). Синаптикалық пластик. Неврологияда (5-ші басылым) (163-182 б.). Сандерленд, Массачусетс: Sinauer Associates.
- ^ Бирн, Дж. (1997). Синаптикалық пластик. Интернеттегі неврологияда (1 бөлім, 7 тарау).
- ^ а б c Bailey, C., Giustetto, M., Huang, Y., Hawkins, R., Kandel, E. (қазан, 2000). Пікірлер: Гетеросинаптикалық модуляция Hebbian пластикасы мен жадысын тұрақтандыру үшін маңызды ма ?. Macmillan Magazines Ltd компаниясында (1-том). Www.nature.com/reviews/neuroscience сайтынан алынды
- ^ а б Хиглей, МДж, Сабатини, Б.Л. (2012 ж. Ақпан.) Дендриттік тікенектердегі кальцийдің сигнализациясы. Биологиядағы суық көктем айлағының болашағы. Алынған http://cshperspectives.cshlp.org/. doi: 10.1101 / cshperspect.a005686.
- ^ а б c г. Редондо, Роджер Л. және Ричард Г.М. Моррис. (2011) «Естеліктерді соңғы рет құру: синаптикалық тегтеу және гипотезаны түсіру». Табиғи шолулар неврология, 12, 17-30.
- ^ Pfeiffer B. E., Huber K. M. (2006). Жергілікті ақуыз синтезі мен синаптикалық пластиканың қазіргі жетістіктері. Неврология журналы, 26 (27), 7147-7150.